
Download
ORIGINAL ARTICLE
Using fruit peel as a substrate for the isolation, biosynthesis, and optimization of glucoamylase by Bacillus sp. under submerged fermentation
Ayesha Kiran1, Memuna Ghafoor Shahid1, Najeeb Ullah2, Abid Sarwar2, Samraz Tahir1, Muhammad Nadeem2, Rubina Nelofer2, Ayaz Ali Khan3, Tariq Aziz4*, Ashwag Shami5, Fahad Al-Asmari6, Fakhria A. Al-Joufi7
1Department of Botany, Government College University, Lahore, Punjab, Pakistan;
2Food and Biotechnology Research Center, PCSIR Labs Complex, Gulberg III, Lahore, Pakistan;
3Department of Biotechnology, University of Malakand Chakdara Dir Lower Khyber Pakhtunkhwa Pakistan;
4Laboratory of Animal Health Food Hygiene and Quality, University of Ioannina – Arta Campus, Arta, Greece;
5Department of Biology, College of Science, Princess Nourah bint Abdulrahman University, P.O. Box 84428, Riyadh 11671, Saudi Arabia;
6Department of Food and Nutrition Sciences, College of Agricultural and Food Sciences, King Faisal University, Al Ahsa, Saudi Arabia;
7Department of Pharmacology, College of Pharmacy, Jouf University, 72341 Aljouf, Saudi Arabia
Abstract
Glucoamylase is a fungal enzyme easily obtained by using a fungal strain—Aspergillus niger. The aim of this study was to isolate the bacterial strain and culture condition optimization to produce glucoamylase. This optimization was achieved under the submerged fermentation technique where fruit peel was used as substrate. The production of glucoamylase was carried out by using (w/v [weight in volume]) yeast extracts 1%, NaCl (chemical formula for sodium chloride) 0.5%, ZnSO4 (zinc sulfate) 0.5%, K2HPO4 (dipotassium phosphate) 0.5, and 1% fruit peel powder as substrate. The optimization of parameters was carried out by one factor at a time technique (OFAT) under submerged fermentation. Different carbon and nitrogen sources were screened for maximum enzyme activity. The physical and chemical parameters such as temperature, pH (potential of hydrogen), inoculum size, and salt concentrations were optimized to achieve the highest production. Based on the biochemical characterizations, the isolated strain was identified as Bacillus sp. FBA1 (fructose-bisphosphate aldolase 1). Maximum production of the enzyme was recorded at the 24th hour at 37°C temperature and pH was kept at 7.0. The salts combinations that produced the highest levels of glucoamylase production from Bacillus strain FBA1 were (g/L) NaCl 1, ZnSO4 1.5, FeSO4 (ferrous sulfate) 2, KH2PO4 (potassium dihydrogen phosphate) 2, CaCl2 (calcium chloride) 0.1, and MgSO4 (magnesium sulfate) 2. The optimum glucoamylase activity of 2.984 mg/mL/minute for FBA1 was obtained at pH 7 at 37°C. The results showed that fruit peels can be exploited as a potential substrate to produce glucoamylase at optimized conditions by Bacillus FBA1.
Key words: Bacillus, fruits peel, submerged fermentation, optimization, glucoamylase
*Corresponding Author: Tariq Aziz, Laboratory of Animal Health Food Hygiene and Quality, University of Ioannina – Arta Campus, Arta, Greece. Email: [email protected]
Academic Editor: Prof. Ilaria Benucci, Department of Agriculture and Forest Sciences (DAFNE), Food and Wine Biotechnology Laboratory, Viterbo, Italy
Received: 5 March 2025; Accepted: 6 June 2025; Published: 1 October 2025
© 2025 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Protease and amylase are the most valuable microbial enzymes, and they have been used extensively and in a variety of applications due to their ability to break down organic waste. which include biofuels, agriculture, pharmaceuticals, chemicals, and biotechnological industries (Ullah et al., 2022). Glucoamylase is an extracellular enzyme that is next to protease in ranking among the industrial enzymes’ distribution. Glucoamylase comes from a variety of sources, including microbial, animals, and plants. Up until the 1970s, enzymes from plants and animals were thought to be considerable, but today enzymes of microbial sources are gaining more popularity (Cherry et al., 2004). Yeast, mold, and bacteria are some of the microorganisms with a higher capacity to produce glucoamylase. The enzymes obtained from these microbial sources are more beneficial than the enzymes obtained from animals and plants (Najafpour et al., 1994). Because of their shorter lifespan, microbial sources are advantageous for the isolation of glucoamylase. Further, enzymes isolated through these sources can easily be obtained and isolated, and their characteristics altered by biotechnology and genetic engineering techniques.
Glucoamylase is a fungal enzyme and easily obtained by using a fungal strain—Aspergillus niger. On an industrial scale, glucoamylase can be produced by the members of genus Aspergillus, Rhizopus, and Bacillus (Bennett, 1998). Although filamentous fungi are a preferred choice to obtain maximum yield on an industrial scale, bacilli bears an edge due to it belonging to a house of thermostable proteins (Bajpai and Bajpai, 1989). Bacilli can also produce enzymes with desirable properties. Almost all species of genus Bacillus have the potential to produce different enzymes (Pretorius et al., 1986). The most important stains of Bacillus widely used for the production of enzymes are: Bacillus licheniformis (Fogarty and Kelly, 1990), B. amyloliquefaciens, B. stearothermophilus (Wind et al., 1994), B. subtilis (Kumar, 2009), and B. megaterium.
Bacteria are an ideal organism for industrial production due to their faster rate of development and simplicity in genetic modification. The cost-effectiveness of the process would increase with the use of thermostable glucoamylase in the starch business due to the possibility of it producing more starch with prolonged enzymatic catalysis (Sauer et al., 2000). The generation of glucoamylase is greatly influenced by the types and concentrations of the carbon and nitrogen sources (Fadel et al., 2020). Behera et al. (2021) studied the impact of adding carbon, nitrogen, and phosphorus on the synthesis of glucoamylase and demonstrated that the medium in solid-state fermentation required the addition of urea, maltose, wheat bran, and potassium biphosphate to produce the most enzymes. The type and concentration of nutrients that are available to the organism affect how an enzyme is produced. Other nutrients may be similarly crucial to the composition of the medium as a sufficient carbon supply. According to Ellaiah et al. (2002), solid-state fermentation can produce glucoamylase using urea as a nitrogen source. According to Behera et al. (2021), adding yeast extract at a very low concentration to a medium containing starch and (NH4)2SO4 (ammonium sulfate) enhances the production of glucoamylase enzyme by A. niger during the submerged fermentation technique. The medium, including starch, wheat bran, (NH4)2SO4, FeSO4 (ferrous sulfate), MgSO4 7H2O (magnesium sulfate heptahydrate), and 0.1N HCl (hydrochloric acid with a normality of 0.1), was said to produce the most glucoamylase.
The induction of the glucoamylase production can be significantly influenced by the pH (potential of hydrogen) and temperature levels (Bertolin et al., 2003; Mishra and Debnath, 2002). Instead of keeping pH at a constant value, there is the possibility of manufacturing glucoamylase under different pH values by using the batch cultivation method. This enzyme was most active at 65°C temperature and 5.0–5.5 pH range (Mamo and Gessesse, 1999), and at 45°C, glucoamylase enzyme produced by A. niger was stable. Nearly 80% of the enzyme’s activity was lost within 75 minutes after the enzyme began to inactivate at 60°C (Ramadas et al., 1996) It turns out that today, glucoamylase is one of the most crucial enzymes in the food industry due to the fact that it is utilized to make glucose syrup from liquid starch (Dale and Langlois, 1940; Nguyen et al., 2002; Nigam and Singh, 1995). Moreover, it is used to produce numerous fermented foods for commercial use in the baking, juice, beverage, pharmaceutical (Fogarty and Kelly, 1990; Hesseltine, 1965), and in some cases, textile, leather, and detergent sectors (Vishnoi et al., 2020).
Food wastes can be used as feedstock in bioproducts to produce enzymes like glucoamylase due to their high protein and carbohydrate content. This is a creative waste management method. In many biotechnological processes, glucose serves as the main carbon source. Nevertheless, depending on where they come from, fruits and vegetables waste are abundant in moisture, carbohydrates, and more. Some fruits and vegetables waste also contain significant amounts of proteins, lipids, and natural colorants, and occasionally antioxidants and other beneficial components, in addition to hydration and carbs. A growing interest in the generation of biochemical from food waste has been observed recently (Lam et al., 2013). Today, submerged fermentation is used for the synthesis of enzymes more all over the world; this method has a lot of potential for producing biocatalysts effectively. It is a low-cost technique that produces a lot of output per volume of the reactor and an easy subject for culture management and downstream operations. Additionally, when crude extracts are applied directly and when the purity of the enzymes is not required, this technique is employed for the production and usage of enzyme (Abdollahi et al., 2021; Shoji et al., 2007). Submerged fermentation has several advantages over traditional solid-state fermentation, one of which is that 90% of all commercial enzymes are extracted from microbes that have been optimized and genetically modified using submerged fermentation (Panpatte and Jhala, 2019)
The development of the genetically engineered microbial strains, the optimization of growth settings, and the choice of suitable microbial strains can all help in increasing and optimizing the production of required enzymes and other primary and secondary metabolites. Further optimization of growth media ingredients helps in improving the effectiveness of the cultivation process (Kumar et al., 2007). The main objective of this study was to isolate glucoamylase producing strain from soil samples and the production of glucoamylase under submerged fermentation using potato peel as substrate.
Material and Methods
Source of bacterial strain
Soil sample for the isolation of bacterial strain for the production of glucoamylase was collected from selected site in PCSIR Laboratories Complex in Lahore, Pakistan.
Isolation of glucoamylase producing Bacillus strain
A 1 g soil sample was weighed and transferred into 9.0 mL sterile saline. Serial dilutions were prepared by transferring 1.0 mL from l0-1 dilution in to separate tubes of up to 10-6. The first four test tubes were discarded, and then the fourth and fifth dilutions were processed under heat shock at 80°C for 15 minutes. This step ensures that if there is any bacterial stain, then it must be of genus Bacillus. Further, 0.1 mL of the sample was transferred to sterilized plates and starch agar medium composition (starch 10 g/L, peptone 5g/L, NaCl [chemical formula for sodium chloride] 2.5 g/L, MgSO4 [magnesium sulfate] 2.5 g/L, and agar 15 g/L) were added. The inoculated plates were incubated at 37°C for 24 hours. After incubation, different colonies appeared, after which the morphologically contrastively colonies were further purified by streaking on nutrient agar medium plates for isolation of pure colonies.
Screening of glucoamylase strain
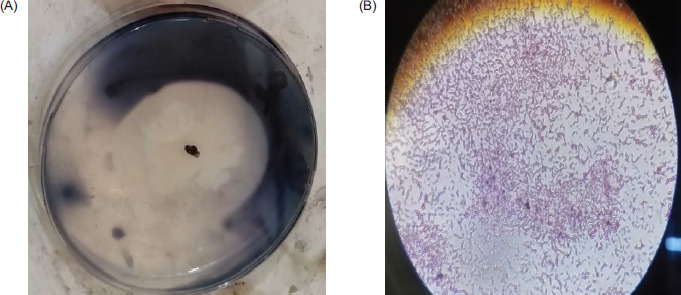
The pure stains were further streaked on the starch agar medium and incubated for 24 hours. Then the iodine solution was flooded on petri plates with bacterial colonies for the identification of bacteria. When iodine reacts with starch, a dark brown color appears. Thus, starch hydrolysis gives a clear zone around the bacterial colony.
Identification of glucoamylase producing strain
Biochemically and morphologically, the strain was identified by the recommended protocol of Bergey’s Manual of Determinative Bacteriology (Holt et al., 1994) and Diagnostic Microbiology (Betty et al., 2002).
AgroIndustrial waste collection
Agroindustrial waste in the form of fruit peels (i.e., apples, bananas, pomegranates, and oranges) were collected as substrate from the vegetable and fruit markets of Lahore.
These peels were dried in an oven for 72 hours and then the dried leaves were ground and the powdered extract of the peel were stored for further use in the production of glucoamylase.
Production medium
The production of glucoamylase was carried out by using (w/v [weight in volume]) yeast extract 1%, NaCl 0.5%, ZnSO4 (zinc sulfate) 0.5%, K2HPO4 (dipotassium phosphate) 0.5%, and 1% fruit peel powder as substrate. The initial pH of the medium was adjusted as 7.0 of the fermentation medium. Initially, the inoculums were added (2% v/v [volume per volume]) and incubated at 37°C for 48 hours. After incubation, the fermented broth was centrifuged at 9000 rpm for 10 minutes with the supernatant used as crude enzyme.
Production of glucoamylase
Different chemicals and physical parameters were optimized. Optimization of fermentation conditions was carried out in a stepwise manner for maximum enzyme production from Bacillus sp. FBA1 (fructose-bisphosphate aldolase 1). Selection of components, including carbon, nitrogen, and mineral components were based in batch fermentation on a one factor at a time (OFAT) methodology.
Screening of carbon and nitrogen sources
Different carbon sources—maltose, galactose, glucose, fructose, and fruit peels—and nitrogen sources—beef extract, peptone, yeast extract, urea, ammonium sulphate, ammonia solution—were screened to obtain suitable carbon and nitrogen source for glucoaylase production.
Optimization of different physiochemical parameters
The optimization of parameters was carried out by the OFAT under submerged fermentation. The effects of various parameters were studied independently.
The effect of various pH on the productivity of glucoamylase enzyme
Various pH such as 3, 4, 5, 6, 7, and 8 were maintained using 1N NaOH (sodium hydroxide solution with a normality of 1) and 1N HCl solution with the help of pH meter in the fermentation medium to observe the maximum yield of glucoamylase enzyme. Comparative optimization was being observed, with each bacterial strain of six replicates.
The effect of various incubation temperature on the productivity of glucoamylase enzyme
Fermentation medium for the optimization of temperature was placed at various incubating temperature—30°C, 35°C, 40°C, and 45°C—for 24 hours in a shaking incubator using submerged fermentation to observe the maximum yield of glucoamylase enzyme. Comparative optimization was being observed, with each bacterial strain of six replicates.
The effect of various inoculum size on the productivity of glucoamylase enzyme
For the optimization of inoculum size, different cementation of inoculum medium—10 mL, 15 mL, 20 mL, and 25 mL—was poured in to the fermentation medium and placed for 24 hours in a shaking incubator using submerged fermentation to observe the maximum yield of glucoamylase enzyme. Comparative optimization was being observed, with each bacterial strain of six replicates.
The effect of various KH2PO4 on the productivity of glucoamylase enzyme
Different concentration of KH2PO4 (potassium dihydrogen phosphate) were measured—0.1, 0.5, 1, 1.5, 2, and 2.5—and added while keeping the other concentration of fermentation ingredients the same, and then placed for 24 hours in a shaking incubator using submerged fermentation to observe the maximum yield of glucoamylase enzyme. Comparative optimization was being observed, with each bacterial strain of six replicates.
The effect of various MgSO4 7H2O on the productivity of glucoamylase enzyme
Different concentration of MgSO4 were measured—0.1, 0.5, 1, 1.5, 2, and 2.5—and added while keeping the other concentration of fermentation ingredients the same and then placed for 24 hours in a shaking incubator using submerged fermentation to observe the maximum yield of glucoamylase enzyme. Comparative optimization was being observed, with each bacterial strain of six replicates.
The effect of various FeSO4 H2O on the productivity of glucoamylase enzyme
Different concentration of FeSO4 H2O (ferrous sulfate monohydrate) was measured—0.1, 0.5, 1, 1.5, 2, and 2.5—and then added while keeping the other concentration of fermentation ingredients same. Fermentation medium were placed for 24 hours in a shaking incubator using submerged fermentation to observe the maximum yield of glucoamylase enzyme. Comparative optimization was being observed, with each bacterial strain of six replicates.
The effect of various CaCl2 on the productivity of glucoamylase enzyme
Different concentration of CaCl2 (calcium chloride) were measured—0.1, 0.5, 1, 1.5, 2, and 2.5—and added while keeping the other concentration of fermentation ingredients same. Then the medium was placed for 24 hours in a shaking incubator using submerged fermentation to observe the maximum yield of glucoamylase enzyme. Comparative optimization was being observed, with each bacterial strain of six replicates.
The effect of the various concentration of NaCl on the productivity of glucoamylase enzyme
Different concentration of NaCl were measured—0.1, 0.5, 1, 1.5, 2, and 2.5—and added while keeping the other concentration of fermentation ingredients the same. Then the medium was placed for 24 hours in a shaking incubator using submerged fermentation to observe the maximum yield of glucoamylase enzyme.
The effect of various ZnSO4 on the productivity of glucoamylase enzyme
Different concentration of ZnSO4 was measured—0.1, 0.5, 1, 1.5, 2, and 2.5—and added while keeping the other concentration of fermentation ingredients same. Then the medium was placed for 24 hours in a shaking incubator using submerged fermentation to observe the maximum yield of glucoamylase enzyme. Comparative optimization was being observed, with each bacterial strain of six replicates.
Analytical procedure
Enzyme assay
A 1 mL supernatant was taken from each flask in to sterilized test tubes with same labelling, and 1 mL starch solution was added in it. The test tubes were placed in an incubator at 37°C for 25 minutes. After incubation, 1 mL of DNS (dextrose normal saline) were added in to each test tube and placed in boiling water for 5 minutes; then 1 mL of distilled water was added in the supernatant samples.
In a 250 mL beaker, 60 mL distilled water was taken, and 1 g DNS was added. This solution was stirred until a clear solution of pale-yellow color was obtained. Then 30 g of potassium sodium titrate was added slowly and stirred well until no crystals remained. Meanwhile, 2N NaOH (sodium hydroxide solution with a normality of 2) was prepared by dissolving 1.6 g of NaOH pellets in 20 mL distilled water. Then, 2N NaOH was added in to the potassium sodium titrate solution and stirred continuously. The final volume was raised to 100 mL and stored in a dark bottle at 4°C.
Statistical analysis
Different parameters were statistically analyzed by one-way ANOVA for comparisons to record the data by using standard error and standard deviation in order to reach the accurate conclusion. Duncan’s multiple range tests were used to compare the significant differences in the mean values at P ≤ 0.05. (Steel and Torrier, 1996).
Results
The two main barriers to the effective application of any technique in the enzyme industry are the overall cost of enzyme manufacturing and downstream processing. Enzymes are typically produced when using any microbial organism with various fermentation procedures, such as solid-state and submerged. The separation of the necessary enzyme is expensive when employing a solid-state fermentation method for glucoamylase synthesis in comparison to a liquid phase system.
Optimization of fermentation condition for maximum glucoamylase production from Bacillus sp
The two main barriers to an effective application of any technique in the enzyme industry are the overall cost of enzyme manufacturing and downstream processing. Enzymes are typically produced when using any microbial organism with various fermentation procedures, such as solid-state and submerged. The separation of the necessary enzyme is more expensive when employing a solid-state fermentation method for glucoamylase synthesis in comparison to a liquid phase system (Imran et al., 2012; Negi and Banerjee, 2009).
Since this process is frequently used for industrial-scale enzyme synthesis and also simple to isolate the required product from the entire fermentation broth, it is frequently utilized when working with bacterial species. Therefore, submerged batch fermentation approach was used for the synthesis of glucoamylase by Bacillus strain FBA1. The fermentation conditions were produced using the OFAT technique.
Effect of various pH on the production of glucoamylase
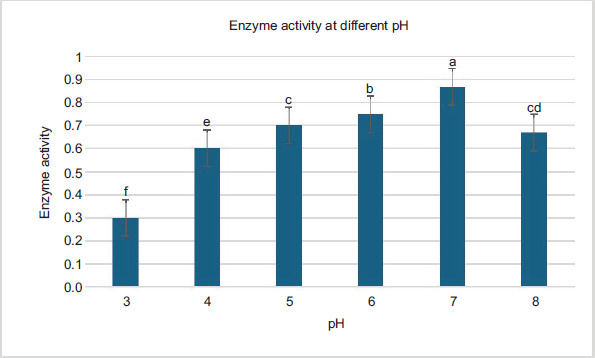
Figure 2 shows the effect of pH on the glucoamylase production from Bacillus sp. The pH of the medium was increased from 3.0 to 8.0. The production of enzyme was observed as maximum (0.073 mg/mL/minute for Bacillus strain) at 7.0 pH. When the pH of the medium was affected from the pH value of 7.0, the yield of the enzyme reduced. At 3.0 pH, the production of the enzyme was low (0.03 mg/mL/minute), and at 8.0 pH, the glucoamylase production was low (0.07 mg/mL/minute). Therefore, optimized pH for the production of enzyme was 7.0 by Bacillus strain.
Figure 1. (A) Iodine test and (B) microscopic identification of Bacillus sp.
Figure 2. Optimization effect of pH.
Effect of incubation temperature
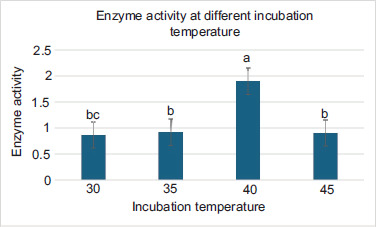
In the production of an enzyme temperature plays an important role. The effect of incubation temperature on glucoamylase synthesis was observed for both Bacillus sp. was incubated in fermentation broth at different temperatures. The Figure 3 depicts the effect of temperature on the production of glucoamylase. A noticeable difference in the production temperature for both the stains was observed. The initial temperature of the medium was varied from 30°C–45°C. Initially, at 30°C the glucoamylase production was extremely low i.e., 0874mg/mL/min (FBA1 strain) When the temperature was raised up to 40°C, the glucoamylase production was also increased. The production of enzyme was observed maximum (1.9mg/mL/min) at 40°C temperature. After this temperature, glucoamylase production decreased due to increase in the temperature and at 45°C the production of the enzyme drastically decreased i.e., 0.913mg/mL. Therefore, 40°C was optimized incubation temperature for the production of glucoamylase enzyme by Bacillus strain.
Figure 3. Optimization effect of temperature.
The development of the bacteria and the synthesis of the enzyme are both significantly influenced by the choice of the optimal temperature. In this study, the impact of incubation temperature on enzyme production by FBA1 strains was examined. It was discovered that the maximal glucoamylase production occurred at 40°C. The synthesis of enzymes ultimately decreased as the incubation temperature was further raised. The biomass eventually ran out to create a substantial amount of enzyme since the growth of the bacteria was temperature-dependent, and a change in the ideal temperature slowed down the growth rate of Bacillus FBA1. The results of other researchers were different from this study—they reported 30°C as optimum incubation temperature for the production of glucoamylase (Mamo and Gessesse, 1999; Mishra and Debnath, 2002).
Effect of various inoculum sizes
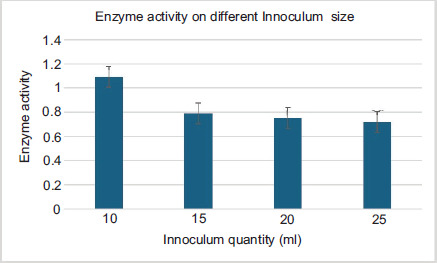
The Figure 4 shows the effect of different inoculum sizes on the glucoamylase production by Bacillus sp. in the shaking fermentation. For inoculation, a 16-hour-old vegetative inoculum was used at different levels 10 mL, 15 mL, 20 mL, and 25 mL (v/v). The enzyme production decreased with an increase in the size of inoculum from 10 mL to 25 mL. At 10 mL, the inoculum size increased with the maximum amount of glucoamylase produced (1.092 mg/mL/minute) after 24 hours.
Figure 4. Optimization effect of inoculum size.
When the size of inoculum was further raised, the production of glucoamylase decreased, although the maximum yield of the enzyme was produced after 24 hours at 10 mL inoculum size. Hence, 10 mL inoculum of Bacillus sp. strain produced maximum amount of the enzyme.
The effect of nitrogen and carbon sources
A medium containing organic nitrogen sources stimulated both the growth of bacteria and enzyme secretion as compared to inorganic nitrogen sources. Of the various nitrogen sources tested in this study, significant growth and enzyme yield was achieved in the medium containing yeast extract (1.955 mg/mL/minute) for Bacillus and almost negligible enzyme production when inorganic nitrogen sources were used (Table 1). All the carbon sources stimulated glucoamylase production in FBA1 stain of Bacillus sp., and fructose proved to be a suitable inducer. The highest amount of enzyme units (1.970 mg/mL/minute) was observed by Bacillus in 24 hours of incubation (Table 1).
Effect of micronutrients
Bacillus strain was tested for the ability to secrete glucoamylase using a variety of micronutrients, including KH2PO4, MgSO4, CaCl2, FeSO4, and NaCl.). Each salt has a unique impact on the generation of enzymes. MgSO4, CaCl2, KH2PO4, FeSO4, ZnSO4, and NaCl concentrations were all altered. The salt combinations that produced the highest levels of glucoamylase secretion from Bacillus strain were (w/v %) KH2PO4 2, NaCl 1, MgSO4.7H2O 2, ZnSO4 1.5, CaCl2 0.1, and FeSO4.7H2O 2 in the production medium (Table 1).
Table 1. Effect of various macro- and micronutrients for the production of glucoamylase.
| KH2PO4 Concentrations (w/v %) | Enzyme Activity mg/ml | min ± SD |
|---|---|---|
| 1 | 0.1 | 0.673±0.0012a |
| 2 | 0.5 | 0.563±0.0082c |
| 3 | 1 | 0.667±0.0053ab |
| 4 | 1.5 | 0.2667±0.0892b |
| 5 | 2 | 0.72±0.0155ac |
| 6 | 2.5 | 0.5273±0.0017b |
| NaCl Concentrations | ||
| 1 | 0.1 | 0.453±0.0155b |
| 2 | 0.5 | 0.545±0.0017ab |
| 3 | 1 | 0.653±0.0082ac |
| 4 | 1.5 | 0.427±0.0055a |
| 5 | 2 | 0.5433±0.002b |
| 6 | 2.5 | 0.533±0.033d |
| MgSO4.7H2O Concentrations | ||
| 1 | 0.1 | 0.295±0.002a |
| 2 | 0.5 | 0.696±0.002c |
| 3 | 1 | 0.637±0.008b |
| 4 | 1.5 | 0.543±0.001528ad |
| 5 | 2 | 0.865±0.002ac |
| 6 | 2.5 | 0.857±0.007b |
| ZnSO4 Concentrations | ||
| 1 | 0.1 | 0.467±0.082a |
| 2 | 0.5 | 0.317±0.019b |
| 3 | 1 | 0.867±0.053d |
| 4 | 1.5 | 1.0317±0.0002c |
| 5 | 2 | 1.023±0.001155d |
| 6 | 2.5 | 0.963±0.067ab |
| CaCl2 Concentrations | ||
| 1 | 0.1 | 1.622±0.055ac |
| 2 | 0.5 | 1.253±0.0089b |
| 3 | 1 | 1.502±0.005cb |
| 4 | 1.5 | 1.56±0.0015b |
| 5 | 2 | 1.307±0.05c |
| 6 | 2.5 | 1.34±0.0115d |
| Nitrogen source | ||
| 1 | Peptone | 1.752±0.0014a |
| 2 | Yeast Extract | 1.955±0.0014b |
| 3 | Urea | 1.747±0.0017c |
| 4 | Beef Extract | 1.624±0.0015ad |
| 5 | Ammonium chloride | 0.426±0.0012ab |
| 6 | Ammonium sulphate | 0.326±0.00066c |
| Carbon source | ||
| 1 | Glucose | 1.514±0.0014a |
| 2 | Fructose | 1.977±0.001ac |
| 3 | Galactose | 1.945±0.0006b |
| 4 | Maltose | 0.347±0.0035a |
| 5 | Sucrose | 0.265±0.0557c |
| 6 | Fruit Peels | 1.47±0.008ab |
| FeSO4.7H2O Concentrations | ||
| 1 | 0.1 | 1.465±0.001a |
| 2 | 0.5 | 1.859±0.0008b |
| 3 | 1 | 2.144±0.001b |
| 4 | 1.5 | 2.043±0.001d |
| 5 | 2 | 2.179±0.001ac |
| 6 | 2.5 | 2.017±0.0011ab |
Discussion
The secretion and stability of the generated enzyme were mostly controlled by the pH of fermentation medium. While the bacterial culture was growing and fermenting, macro- and micronutrients were being consumed, and numerous metabolites or byproducts were created along with the required enzyme. The pH change was ultimately caused by all these components. Depending on the type of microbial source employed, glucoamylase can be generated in both acidic and basic conditions (Fadel et al., 2020). According to Nigam and Singh (1995), LactoBacillus amylovorus produced the most glucoamylase at pH 6.0. However, archaeal species like Thermoplasma acidophilum, Picrophilus torridus, and P. oshimae are also among the stains that were modified to produce enzyme at very high acidic pH levels (Adhiyanto et al., 2021).
The development of the bacteria and the synthesis of the enzyme are both significantly influenced by the choice of the optimal temperature. In this study, the impact of incubation temperature on enzyme production FBA1 strains was examined, and the result showed that the maximal glucoamylase production occurred at 40°C. The synthesis of enzymes ultimately decreased as the incubation temperature was further raised. The biomass eventually ran out to create a substantial amount of enzyme since the growth of the bacteria was temperature-dependent, and a change in the ideal temperature slowed down the growth rate of Bacillus FBA1. The results from other researchers were different from this study, as they reported 30°C as optimum incubation temperature for the production of glucoamylase (Mamo and Gessesse, 1999; Mishra and Debnath, 2002).
Fogarty and Kelly (1990) isolated glucoamylase from C. themohdroysulfuricum and studied the effect of different carbon sources on its production. From their findings, it was concluded that the soluble starch from potato showed a comparatively high titer of glucoamylase. Furthermore, Kumar et al. (2007) also enhanced glucoamylase production using a more increased concentration of soluble starch (5.0%). Other researchers have also supplemented the growth medium with a variety of starch and varied the concentration to further enhance the production of enzyme (Kumar, 2009). Similarly, Arifeen et al. (2024) isolated the amylase producing the Bacillus ichenoformis strain from garden soil of Lichi chinesis and used rice polish as substrate. Helal et al. (2021) reported that when peptone is added in combination with the yeast extract, optimum enzyme was produced by Rhizopus sp., whereas when these two components were used separately, they produced comparatively less amount of enzyme. Similarly, Helal et al. (2021) also used a mixture of peptone and yeast extract for the optimum production of enzyme. However, when both these complex nitrogenous sources were used they induced excessive proteolytic enzyme secretion which ultimately leads to the hydrolysis of various amylotic enzymes (Vishnoi et al., 2020).
The production of enzymes is mostly Ca+2 (calcium ions) dependent because most of the α-amylase enzymes are considered as metalloenzymes. The studies reveal that when CaCl2 is added in the production media, a higher yield of enzymes were observed (Zare et al., 2021). In this study, different concentrations of CaCl2 (0.1 g, 0.5 g, 1 g, 1.5 g, 2 g, 2.5 g) were added in the production media, and a maximum of glucoamylase production was observed in 0.1 g for Bacillus. Similar results were also reported by Kumar et al. (2007) for the production of thermostable α-amylase from Bacillus subtilis KIBGE HAR—a newly isolated strain.
An increase in the production of enzymes might be due to the fact that calcium plays a vital role in the physiology and metabolism of bacteria. It also provides stability to the desired enzyme against the proteases, which was also produced during the process of fermentation (Manfredini et al., 2021). When the concentration of calcium increased more than 0.1 grams, decline in the enzyme production was clearly observed. It could be because of bacterial growth that ultimately affect the production of enzyme (Manfredini et al., 2021).
Conclusion
The aim of this study was to isolate an economical source for the production of glucoamylase enzyme that could be industrially desirable. Glucoamylase, a cost-effective enzyme, was produced from Bacillus sp. using the submerged fermentation. Fruit peels were used as substrate as they are budget- and eco-friendly carbon source for the production of enzyme. The maximum production of glucoamylase was obtained from a cost-effective thermostable Bacillus FBA1 strain. The optimum pH and temperature for the production of glucoamylase from Bacillus FBA1 were 7.0 and 37°C. Ideal incubation period for the growth of bacteria were 24 hours with a 10 mL inoculum size. Glucoamylase activity were 2.984 mg/mL/minute with 5 g fruit peel as a carbon source and yeast as nitrogen source for 24 hours at pH 7 and 37°C temperature.
Data Availability Statement
All the data generated in this research work has been included in the manuscript.
Acknowledgments
Authors are thankful to the Princess Nourah bint Abdulrahman University Researchers Supporting Project Number PNURSP2025R31, Princess Nourah bint Abdulrahman University, Riyadh, Saudi Arabia. The authors also express their gratitude to the Deanship of Scientific Research (DSR) at King Faisal University under the Project no. [KFU250822].
Author Contributions
Conceptualization: Memuna Ghafoor Shahid; methodology: Ayesha Kiran; software: Najeeb Ullah; validation: Abid Sarwar; formal analysis: Samraz Tahir; investigation: Muhammad Nadeem; resources: Rubina Nelofer; data curation: Fahad Al-Asmari; writing—original draft preparation: Ayesha Kiran; writing—review and editing: Fakhria A. Al-Joufi; visualization: Ashwag Shami; supervision: Tariq Aziz; project administration: Ayaz Ali Khan; funding acquisition: Tariq Aziz.
Conflicts of Interest
The authors declare no conflicts of interest.
REFERENCES
Abdollahi F, Jahadi M, Ghavami M. Thermal stability of natural pigments produced by Monascus purpureus in submerged fermentation. Food Sci Nutr. 2021;9(9):4855–62. 10.1002/fsn3.2425
Adhiyanto C, Rahmawati NM, Saibi Y, Mutia Nasir N, Hendarto H, Sari FR, et al. Identification of the Archaeon Picrophilus torridus isolated and cultured from Tangkuban Perahu and Dieng Plateau on Java Island, Indonesia. Copyright@ EM International. 2021;27(3):1186–95.
Arifeen S, Jamil J, Sarwar A, Ullah N, Nelofer R, Aziz T, et al. Biosynthesis and optimization of amylase from bacillus sp. Isolated from soil samples using agro-industrial waste as substrate. Appl Ecol Environ Res. 2024;22(4):2927–40.
Bajpai P, Bajpai PK. High-temperature alkaline a-amylase from Bacillus licheniformis TCRDC-B13. Biotech Bioeng. 1989;
Behera BC, Mishra R, Mohapatra S. Microbial citric acid: Production, properties, application, and future perspectives. Food Front. 2021;2(1):62–76. 10.1002/fft2.66
Bennett JW. Mycotechnology: The role of fungi in biotechnology. J Biotech. 1998;66(2–3):101–7. 10.1016/S0168-1656(98)00133-3
Bertolin TE, Schmidell W, Maiorano AE, Casara J, Costa JAV. Influence of carbon, nitrogen and phosphorous sources on glucoamylase production by Aspergillus awamori in solid state fermentation. ZNC J Biosci. 2003;58(9–10):708–12. 10.1515/znc-2003-9-1020
Betty AF, Daniel FS, Alice SW. Overview of bacterial identification methods and strategies. In: Bailey and Scott’s Diagnostic Microbiology. 11th ed. Mosby Inc.: USA, 2002. 225–83 pp. 10.1093/clinchem/48.10.1816
Cherry HM, Hossain MT, Anwar MN. Extracellular glucoamylase from the isolate aspergillus fumigatus. Pak J Biol Sci. 2004;7(11):1988–92. 10.3923/pjbs.2004.1988.1992
Dale JK, Langlois DP. Sirup and method of making the same. United States Patent, 2: 201, 609. 1940:2–7.
Ellaiah P, Adinarayana K, Bhavani Y, Padmaja P, Srinivasulu B. Optimization of process parameters for glucoamylase production under solid state fermentation by a newly isolated Aspergillus species. Process Biochem. 2002;38(4):615–20. 10.1016/S0032-9592(02)00188-7
Fadel M, AbdEl-Halim S, Sharada H, Yehia A, Ammar M. Production of glucoamylase, α-amylase and cellulase by Aspergillus oryzae F-923 cultivated on wheat bran under solid state fermentation. J Adv Biol Biotechnol. 2020:8–22. 10.9734/jabb/2020/v23i430149
Fogarty WM, Kelly CT. Recent advances in microbial amylases. EMT. 1990:71–132. 10.1007/978-94-009-0765-2_3
Helal SE, Abdelhady HM, Abou-Taleb KA, Hassan MG, Amer MM. Lipase from Rhizopus oryzae R1: In-depth characterization, immobilization, and evaluation in biodiesel production. J Genet Eng Biotechnol. 2021;19(1):1–13. 10.1186/s43141-020-00094-y
Hesseltine CW. A millennium of fungi, food, and fermentation. Mycologia. 1965;57(2):149–97. 10.1080/00275514.1965.12018201
Holt JG, Krieg NR, Sneath PHA, Stately JT, William ST. Bergey’s manual of determinative bacteriology, 9th edn. Williams and Wilkins: Baltimore, USA, 1994. 559 p.
Imran M, Asad M, Gulfraz M, Qureshi R, Manzoor N, Choudhary A. Glucoamylase production from Aspergillus niger by using solid state fermentation process. Pakistan Journal of Botany. 2012;44(6):2103–10.
Kumar GS. Enhanced production and partial purification of glucoamylase from mutated Bacillus sp. FME. J Korean Soc Appl Biol Chem. 2009;52(5):412–8. 10.3839/jksabc.2009.073
Kumar S, Kumar P, Satyanarayana T. Production of raw starch-saccharifying thermostable and neutral glucoamylase by the thermophilic mold Thermomucor indicae-seudaticae in submerged fermentation. Appl Biochem Biotechnol. 2007;142(3):221–30. 10.1007/s12010-007-0011-x
Lam WC, Pleissner D, Lin CSK. Production of fungal glucoamylase for glucose production from food waste. Biomolecules. 2013;3(3):651–61. 10.3390/biom3030651
Mamo G, Gessesse A. Production of raw-starch digesting amyloglucosidase by Aspergillus sp. GP-21 in solid state fermentation. J Ind Microbiol Biotechnol. 1999;22(6):622–6. 10.1038/sj.jim.2900676
Manfredini PG, Cavanhi VAF, Costa JAV, Colla LM. Bioactive peptides and proteases: characteristics, applications and the simultaneous production in solid-state fermentation. Biocatalysis and Biotransformation. 2021;39(5):360–77. 10.1080/10242422.2020.1849151
Mishra A, Debnath M. Effect of pH on simultaneous saccharification and isomerization by glucoamylase and glucose isomerase. Appl Biochem Biotechnol–Part A Enzyme Engineering and Biotechnology. 2002:102–3, 193–9. 10.1385/ABAB:102-103:1-6:193
Najafpour GD, Klasson KT, Ackerson MD, Clausen EC, Gaddy JL. Biological conversion of poultry processing waste to single cell protein. Bioresour Technol. 1994;48(1):65–70. 10.1016/0960-8524(94)90138-4
Negi, S., Banerjee, R. Optimization of extraction and purification of glucoamylase produced by Aspergillus awamori in solid-state fermentation. Biotechnol Bioproc E. 2009;14: 60–66. 10.1007/s12257-008-0107-3
Nguyen QD, Rezessy-Szabó JM, Claeyssens M, Stals I, Hoschke Á. Purification and characterisation of amylolytic enzymes from thermophilic fungus Thermomyces lanuginosus strain ATCC 34626. Enzyme and Microbial Technology. 2002;31(3):345–52. 10.1016/S0141-0229(02)00128-X
Nigam P, Singh D. Enzyme and microbial systems involved in starch processing. Enzyme Microb Technol. 1995;17(9):770–78. 10.1016/0141-0229(94)00003-A
Panpatte DG, Jhala YK. Soil fertility management for sustainable development. In: Soil fertility management for sustainable development (Issue October). Springer: Singapore, 2019. 10.1007/978-981-13-5904-0
Pretorius IS, Kock MJD, Britz TJ, Potgieter HJ, Lategan PM. Numerical taxonomy of α-amylase producing Bacillus species. J Appl Bacteriol. 1986;60(4):351–60. 10.1111/j.1365-2672.1986.tb01743.x
Ramadas M, Holst O, Mattiasson B. Production of amyloglucosidase by Aspergillus niger under different cultivation regimens. World J. Microbiol Biotechnol. 1996;12(3):267–71. 10.1007/BF00360926
Sauer J, Sigurskjold BW, Christensen U, Frandsen TP, Mirgorodskaya E, Harrison M, et al. Glucoamylase : Structure/function relationships and protein engineering. PubMed. 2000;1543(2):275–93.
Shoji H, Sugimoto T, Hosoi K, Shibata K, Tanabe M, Kawatsura K. Simultaneous production of glucoamylase and acid-stable α-amylase using novel submerged culture of Aspergillus kawachii NBRC4308. J Biosci Bioeng. 2007;103(2):203–5. 10.1263/jbb.103.203
Steel, R., Torrie, J. and Dickey, D. (1996) Principles and Procedures of Statistics: A Biometrical Approach. 3 Sub Edition, McGraw-Hill, New York.
Ullah N, Rehman MU, Sarwar A, Nadeem M, Nelofer R, Shaki, HA, et al. Purification, characterization, and application of alkaline protease enzyme from a locally isolated Bacillus cereus strain. Fermentation. 2022;8(11):628.
Vishnoi N, Dixit S, Mishra J. Microbial lipases and their versatile applications. Springer: Singapore, 2020. 10.1007/978-981-15-1710-5_8
Wind RD, Buitelaar RM, Eggink G, Huizing HJ, Dijkhuizen L. Characterization of a new Bacillus stearothermophilus isolate: A highly thermostable α-amylase-producing strain. Appl Microbiol Biotechnol. 1994;41(2):155–62. 10.1007/BF00186953
Zare H, Meiguni F, Najafpour GD. Production of alkaline protease using industrial waste effluent as low-cost fermentation substrate. Iranian Journal of Energy and Environment. 2021;12(3):264–72. 10.5829/ijee.2021.12.03.11