
Download




PAPER
Evaluation of the durum wheat landrace genetic diversity using agro analysis and its benefit for human health
Mohammad Ihsan1, Arshad Khan1, Nausheen Nazir2, Mohammad Nisar1*, Tour Jan1, Shariat Ullah1, Tariq Aziz3*, Fahad Al-Asmari4, Sahar A. Alshareef5, Maha Aljabri6, Manal Y. Sameeh7
1Department of Botany, University of Malakand, Chakdara 18800 Pakistan;
2Department of Biochemistry, University of Malakand, Chakdara 18800 Pakistan;
3Laboratory of Animal Health, Food Hygiene, and Quality, Department of Agriculture, University of Ioannina, 47100 Arta, Greece;
4Department of Food and Nutrition Sciences, College of Agricultural and Food Sciences, King Faisal University, Saudi Arabia;
5Department of Biology, College of Science and Arts at Khulis, University of Jeddah, Jeddah, Saudi Arabia;
6Department of Biology, Faculty of Sciences, Umm Al-Qura University, Makkah, Saudi Arabia;
7Department of Chemistry, Al-Leith University College, Umm Al-Qura University, Makkah, Saudi Arabia
Abstract
Ancient wheat and landraces (traditional varieties) are fascinating to the consideration of scientists who are re-evaluating the healthy and dietary properties attributed to them by popular tradition. Wheat (Triticum durum) is one of the most significant crops that is grown all over the world. In the present study, 19 landraces were analyzed for genetic diversity through morphological and SDS-PAGE analysis. The highest coefficient of variance (18.91%) was found for spike length, and the lowest for days to maturity (1.51%). The genotype Malakand and Dolram produced a maximum number of seeds (42 and 46 seeds per plant), and the genotypes Dogai and Rasool Banda were found to be the tallest (73 and 71 cm, respectively). The genotype Kamar Kotki was found to mature early (165 days) while Doagi was found to mature late (171 days). Spike length was found to be positively correlated with plant height, leaf width, and leaf length. Leaf width was significant with spike length and the internode distance. The cluster analysis divided all the genotypes into six sub-clusters where the Asban and the Shekhan were placed at the extreme of the dendrogram. Based on the total seed storage proteins analysis, 18 (85.71%) bands were polymorphic while 14.28% bands were found to be monomorphic. Using Ward’s methods, all the genotypes were divided into two main lineages at 25% Euclidian distance and five sub-clusters at 75% distance. Our findings have significance for understanding the variations and relationships between various durum wheat genotypes and could serve as a basis for local elite durum germplasm that has been regionally adapted.
Key words: Genetic diversity, Durum wheat, morphology, genotypes, SDS-PAGE
Corresponding Authors: Mohammad Nisar, Department of Botany, University of Malakand, Chakdara 18800 -Pakistan. Email: [email protected]; Tariq Aziz, Laboratory of Animal Health, Food Hygiene, and Quality, Department of Agriculture, University of Ioannina, 47100 Arta, Greece. Email: [email protected]
Received: 29 October 2023; Accepted: 12 December 2023; Published: 2 January 2024
© 2024 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Triticum durum (2n=4×=28, with AABB genotype), a member of the Poaceae family, is an important source of food products like pasta, noodles, and macaroni (Zhang & Cai 2019). It has been the main staple in Europe, North Africa, and Central Asia for 8,000 years, and it is the leading cereal grain in terms of harvested area and nutrition. It provides around 19% of the world’s population with calories and 21% of their protein needs. Only 5% of the wheat produced worldwide is tetraploid (durum wheat), according to a review of the research that is currently accessible (Niaz et al., 2014).
Durum wheat always gets a more favorable price on the market than bread wheat, because of its commercial significance in the production of bakery goods. It is important to assess the genetic diversity already present in the germplasm for the improvement of durum wheat (like any other crop species) (Khan et al., 2022; Awulachew, 2020). Having the greatest area than any other crop species, wheat is grown on more than 240 million hectares. The bread wheat does exhibit a range of phenological responses to a wide range of environmental variables due to the genotype shaped by the polyploidy and vast genetic variability (Feuillet et al., 2008). Variation among plant species at the genetic level is an essential part of any productive breeding program. As a result, genetic diversity assessment is essential for the efficient use of genetic resources in breeding programs (Khan et al., 2015). In addition, in other breeding programs for wheat improvement, variations in the traits are important in the field of diversity (Khodadadi et al., 2011). To choose the best parents, it is crucial to determine the correlations between genotypes (Ghasemi et al., 2019). According to Khaled et al., (2015), approaches for identifying and evaluating the genotypes have evolved from the examination of various discrete characteristics. The development of breeding populations and the application of selection procedures may benefit from the characterization of wheat genotypes (Aghaee et al., 2010). In addition, the identification of plant genomes and classification of the genes responsible for significant features have been accomplished for the molecular markers study (Khan et al., 2015). As a result, morphological traits were successfully utilized to assess the genetic variety in wheat (EL-Rawy and Youssef, 2016). Therefore, the use of molecular markers for assessment of the genetic diversity receives much attention from wheat breeders (Salem et al., 2015).
By application of morphological traits and molecular markers, wheat genetic diversity has been thoroughly assessed. Genetic diversity analysis has been effectively applied to phenotypic identification based on physical features (Zarkti et al., 2010). Poor polymorphism, poor heritability, late expression, and the possibility of epistatic and pleiotropic gene effects are only a few of the drawbacks of morphological features (Kesralikar et al., 2017). On the contrary, protein markers are useful tools for cultivar identification, new variety registration, and overall crop classification as well as for studying genetic diversity, which increases the effectiveness of wheat breeding programs in cultivar development (Varzakas et al., 2014). Seed storage proteins, for example, reflect genotypes more accurately and independently of environmental effects than do protein markers (Shuaib et al., 2007). Thus, the present study aimed to assess the genetic diversity of durum wheat genotypes using different morphological traits and estimation of seed protein polymorphism using SDS-PAGE analysis.
Materials and Methods
Plant material
The present research study was conducted at the Department of Botany, Botanical Garden and Herbarium, University of Malakand KPK Pakistan, between 2020 and 2021. A total of 19 genotypes of durum wheat (Triticum durum) were collected from different regions of Malakand and evaluated for phenotypic and genetic traits (Table S1). Plants were sown on 15 November 2020 in randomized complete block design (RCBD). In the field, the row-to-row distance was 45 cm and the row length was 4 m.
Morphological characterization
During the present study, nine quantitative traits were recorded. The data obtained from these traits were collected from five guarded plants and their mean value was used for data analysis. The quantitative traits were days to germination, days to flowering, days to maturity, plant height, leaf length, leaf width, spike length, awns length, node distance, and the number of seeds per plant. Days to germination were recorded from the date of sowing when the plant was germinated, and the days were counted. Days to flowering were noted for all plants in each row from the sowing date when the first flower was initiated in the germplasm. Days to maturity were noted when the plants began to turn yellow. Plant height was recorded when the plant reached maturity stage. Leaf length was measured from each randomly selected genotype and measurement was carried out from the base to the tip of each leaf with the help of a meter rod. For the width of the leaf, five leaves were randomly selected from each genotype, and measurement was carried out with the help of a meter rod. For the length of the spike, five spikes were randomly selected from each genotype and measurement was carried out from the base to the tip of each spike with the help of a scale. Awn lengths were randomly selected from each genotype and measurement was carried out from the base to the tip of each awn with the help of a meter rod scale. Node distance was measured in each line and their mean value was used for data analysis. Seeds per plant were counted, and their mean value was used for statistical packages.
Evaluation of the protein polymorphism
The seeds of all 19 genotypes were subjected to SDS-PAGE analysis to determine the genetic diversity among the studied genotypes. Healthy and mature seeds from each genotype were finely ground with mortar and pistil. About 0.02 g of the powder seeds were transferred to 1.5 mL Eppendorf tubes. After that, 400-μl protein extraction buffer (Distilled water 70 mL, Tris 0.60 g, SDS 0.2 g, Urea 30.3 g, 2-Mercaptoethanol 1mL, and pH 8) was added to each tube. The powder was then homogenized at room temperature for 2 min before centrifugation at 13,000 rpms (Revolutions Per Minute) for 15 minutes. About 12.25% polyacrylamide gel was used to make SDS-PAGE slabs, consisting of two phases of separation gel and stacking gel. The composition of the separation gel was: 20% acrylamide with 0.135% N.N-methylene-acrylamide in 0.15 M Tris-HCl buffer with a pH of 8.8 and SDS of 0.27%. For polymerization of the gel, 10% APS (Ammonium per sulfate) and 15 microliters of Tetramethylethylene-diamine (TEMED) were added to the solution. For the making of the stacking gel, 0.8% N.N-methylene-bis-acryalmide, 30% acrylamide 0.25 Tris (pH of 6.8), 0.2% SDS, 70 µl APS, and 17 µl TEMED were mixed. About 10 µl of the sample solution was loaded into the well of the stacking gel through a micropipette. The gel plates were kept in the gel tank filled with electrode buffer solution (0.025M Tris, 0.129M Glycine, 0.125% SDS). About 100 voltage current was applied to get the electrophoretogram. When the blue line of bromophenol blue reached the bottom of the gel, the electric current supply was disconnected. After that, the gel plates were taken out from the assembly and transferred to a plate filled with staining solution (Distilled water 500 mL, Acetic acid 60 mL, Methanol 440 mL, Coomassie brilliant blue (CBB)* R250 2.25 g) and gently rotated on a shaker for 2 h. After that, for de-staining (Distilled water 750 mL, Methanol 220 mL, and Acetic acid 50 mL), gels were kept in the destining buffer for several hours overnight till the disappearance of the blue color for the gel and protein subunits were observed (Nisar et al., 2009)
Data analysis
For the estimation of genetic diversity, all the recorded data of morphological traits obtained from the 19 wheat genotypes were averaged, and the mean value was subjected to different statistical packages. Basic statistics (mean, maximum, minimum, standard error, standard deviation, and CV%) were computed using MS Excel 2016. The correlation coefficient was computed using SPSS software version 22 and cluster analysis, and the principal component analysis (PCA) was performed using PC ORD software version 5 (Wagga Wagga, Australia). Depending on the presence and absence of polypeptide protein sub-units, similarity index was planned for the total possible pairs of different protein types. Similarly, for total seed storage proteins, the binary data matrices were analyzed via unweighted pair group arithmetic average (UPGMA), genetic similarity analysis, and genetic linkage analysis. For data scoring, ‘1’ was used for band presence while null (‘0’) was used for band absence. The binary data were then subjected to cluster analysis, A two-way cluster dendrogram was constructed using PC-ORD software version 5 (Wagga Wagga, Australia).
Results
Diversity in morphological characterizations
Descriptive statistics (mean, maximum, minimum, standard error, standard deviation, and coefficient of variation) of different morphological characters are presented in Table 1. In the present study, significant variation was found for most of the morphological traits, the coefficient of variation percent (CV%) ranged from 1.5 to 18.9%, with the highest CV% for spike length (18.91%), awn length (18.32%), and leaf length (17.90%), whereas the lowest coefficient of variation percent was recorded for days to maturity (1.51%) and days to flowering (2.07%). During the present investigation, a significant level of variation was observed for day to germination, ranging from 10 to 14 days with a CV% of 9.38%; the early germinated genotype was Tangi Dara which took 10 days to germinate, followed by Kolal Derai with 11 days, while the late germinated genotypes were Malakand and Dogai which took 14 days to germinate.
Table 1. Descriptive statistics of different quantitative traits used in the present study.
| Traits | Minimum | Maximum | Mean | Standard Error | Standard Deviation | CV% |
|---|---|---|---|---|---|---|
| Days to germination | 10 | 14 | 12.37 | 0.27 | 1.16 | 9.38 |
| Days to flowering | 107 | 114 | 110.42 | 0.53 | 2.29 | 2.07 |
| Days to maturity | 165 | 171 | 166.26 | 0.58 | 2.51 | 1.51 |
| Plant height | 46.75 | 73.23 | 62.84 | 1.68 | 7.3 | 11.62 |
| Leaf length | 10 | 24.75 | 18.1 | 0.74 | 3.24 | 17.90 |
| Spike length | 5.75 | 11.63 | 8.62 | 0.37 | 1.63 | 18.91 |
| Leaf width | 0.83 | 1.38 | 1.14 | 0.03 | 0.15 | 13.16 |
| Node distance | 14.25 | 19.38 | 16.69 | 0.37 | 1.63 | 9.77 |
| Awn length | 4.63 | 9.63 | 7.15 | 0.3 | 1.31 | 18.32 |
| Number of seed/plant | 29 | 46.25 | 38 | 0.92 | 4.02 | 10.58 |
For days to flowering, the data ranged from 107 to 114 days with a mean value of 110.42 and a coefficient of variation percent of 2.07%. The early days to flowering genotypes were Tangi Dara and Negotal which took 107 days to reach the stage of flowering, followed by Sandai with 111 days, and 113 days to flowering was observed in many genotypes, namely, Asbanr Dogai and Rasool Banda while the late flowering genotype was Rabat which took 114 days to reach the stage of flowering.
Analysis of variance revealed significant variation for days to maturity, where the data ranged from 165 to 171 days with a mean value of 166.26 and coefficient of variation percent of 1.51. The early days to maturity (165) were observed for genotypes Tangi Dara, Daroo, Kolal Derai Hayaserai, Negotal, Sarpato and Malakand while the late mature genotypes were Dogai, Rasool, Banda, Dol Ram, and Matta Swat with 171 days to reach the stage of maturity.
Plant height ranged from 46.75 to 73.23 cm with a mean value of 62.84 cm and a coefficient of variation percent of 11.61. The minimum plant height (46.75) cm was observed for genotype ‘Sheikhano’ followed by Matta Swat, which had a 55 cm plant height, whereas the tallest genotype was Dogai, which had a 73.23 cm plant height followed by Rasool Banda having a plant height of 71.62.
Similarly, leaf length ranged from 10 to 24.75 cm with a mean of 18.1 cm and a coefficient of variation percent of 17.90%. The maximum leaf length of 24.75 cm was recorded for Rabat followed by Asban. The lowest leaf length (10) cm was recorded for Sarpato and Matta Swat had a leaf length of 15 cm.
The spike length ranged from 5.75 to 11.63 cm with a mean of 8.62 and a coefficient of variation percent of 18.91. The minimum spike length of 5.75 cm was recorded for Sarpato followed by Sheikhano having a spike length of 5.87 cm while Jannat Banda had a spike length of 5.97 cm, whereas the maximum spike length (11.63) cm was recorded for the Sandai genotype.
The leaf width ranged from 0.83 to 1.38 cm with a mean of 1.14 cm, and a coefficient of variation percent of 13.16, where the minimum leaf width (0.83) was recorded for Sarpato and the maximum 1.38 was found for Negotal.
The node distance ranged from 14.25 to 19.38 with a mean of 16.69, and a coefficient of variation percent of 13.16. The maximum node distance of 19.38 cm was recorded for Dogai followed by Tan I Dara at 18.62 cm and the minimum of 14.25 cm was recorded for Sarpato followed by Sadoo with a node distance of 14.37 cm.
Furthermore, the awn length was recorded with a range of 4.63 to 9.63 with a mean of 7.15 cm and a coefficient of variation percent of 18.32. The maximum awn length of 9.63 cm was recorded for Dogai and Hayaserai and the minimum of 4.63 cm was found for Sarpato followed by Malakand having 6.75 cm awn length.
The number of seeds per plant ranged from 29 to 46.25 g with a mean of 38 g and a coefficient of variation percent of 10.58. The minimum number of seeds per plant (29) was recorded for Jannat Banda, followed by Hayaseri having 34 seeds per plant, while the maximum of 46.25 g was recorded for Dol ram followed by Asban and Kamar Kotki, respectively.
Correlation analysis
Correlation analysis was computed for different quantitative parameters using SPSS software version 22 (Table 2). In the current study, it was found that plant height was significantly correlated with days to maturity (r = 0.35*), spike length was significantly correlated with plant height (r = 0.73**), and leaf length (r = 0.64**) and leaf width were found to be significantly correlated with leaf length (r = 0.50*). Node distance was found to be significantly correlated with plant height (r = 072**) and with spike length (r = 0.69**).
Table 2. Correlation coefficient analysis of quantitative traits of durum genotype.
| Traits | DG | DF | DM | PH | LL | SPKL | LW | NDS | AWL | NSP |
|---|---|---|---|---|---|---|---|---|---|---|
| DF | 0.10 | 1 | ||||||||
| DM | 0.17 | 0.36 | 1 | |||||||
| PH | 0.23 | 0.37 | 0.35* | 1 | ||||||
| LL | -0.01 | 0.13 | -0.29 | 0.35 | 1 | |||||
| SPKL | 0.11 | 0.29 | 0.19 | 0.73** | 0.64** | 1 | ||||
| LW | 0.04 | -0.08 | -0.13 | 0.22 | 0.50* | 0.73** | 1 | |||
| NDS | 0.08 | 0.29* | 0.28* | 0.72** | 0.42 | 0.69** | 0.32 | 1 | ||
| AWL | 0.20 | 0.16 | 0.26 | 0.26 | 0.35* | 0.37 | 0.23 | 0.37 | 1 | |
| NSP | 0.04 | -0.05 | 0.27 | 0.37* | -0.07 | 0.24 | -0.01 | 0.37 | -0.26 | 1 |
**Correlation is significant at the 0.01 level
*Correlation is significant at the 0.05 level
Note: DG-Days to germination, DF-Days to flowering, DM-Days to maturity, PH-Plant height, LL-Leaf length, SPKL-Spike length, LW-Leaf width, NDS-Node distance, AWL-Awn length, NSP-Number of seed -1.Supplementary
Cluster analysis
Ward’s method was used for cluster analysis using PC ORD software ver.6. The cluster dendrogram was constructed using different quantitative traits. A 25% Euclidian distance was used to divide all the genotypes into two lineages, and a 75% Euclidian distance was used to further subdivide them into six clusters (Figure 1). Cluster 1 consisted of four genotypes (Asban, Rabat, Sadai, Tangi dara), Cluster 2 comprised five genotypes (Daro, Nagotal Shai, Hayseri, Sadoo), sub-cluster 3 had three genotypes (Dogai, Rasool, Dalram). Cluster 4 consisted of five genotypes (Kamarkotki, Malakand, Kolal Deri, Sarpato, Matta Swat). Cluster 5 had one genotype Jannat Banda and the last sub-cluster also had one genotype Shekahnu.
Figure 1. Cluster dendrogram of different quantitative traits of durum wheat used in the present study.
Principal component analysis (PCA)
PCA is helpful for the selection of the best cultivar among the studied genotypes. Among the studied genotypes, all the PCs, (PC1, PC2, PC3, and PC4) show 78.08% of the total variation in the data set (Table 3, Figure 2). To plan for the utilization of suitable gene pools in crop development for specific plant traits, PCA aids in the conservation and use of genetic resources (Zubair et al., 2007).
Table 3. Principal component study of 19 durum wheat genotypes collected from the Malakand region for quantitative traits.
| AXIS | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| Cumulative variance (%) | 35.08 | 53.93 | 68.91 | 78.08 |
| Days to germination | -0.13 | 0.10 | 0.20 | 0.84 |
| Days to flowering | 0.19 | 0.01 | 0.47 | 0.34 |
| Days to maturity | -0.17 | 0.33 | 0.47 | -0.03 |
| Plant height | -0.43 | 0.09 | 0.12 | -0.13 |
| Leaf length | -0.30 | 0.41 | -0.22 | -0.06 |
| Spike length | -0.47 | 0.16 | -0.09 | -0.06 |
| Leaf width | -0.29 | 0.31 | -0.36 | 0.16 |
| Node distance | -0.43 | 0.01 | 0.06 | -0.18 |
| Awn length | -0.22 | 0.33 | 0.36 | 0.24 |
| Number of seed/plant | -0.21 | 0.54 | -0.27 | -0.08 |
Figure 2. Scatter plot of quantitative traits of durum wheat genotypes collected from the Malakand region.
In the first principal component, the total variation was 35.08%, and they were associated positively with days to flowering, while the days to germination, days to maturity, plant height, leaf length, leaf width, nodes distance, and number of seeds per plant contributed negatively with the PC1.
The second principal component accounted for a total variation of 53.93%, and it was found that the days to germination, days to flowering, days to maturity, plant height, leaf length and width, length of the awns, and the number of seeds per plant were found to positively contribute toward PC2.
The overall variation in PC3 was 68.91%, and it was found that the contribution of leaf length, leaf width, and number of seeds per plant negatively contributed while days to flowering, days to maturity, plant height, and awn length positively contributed in PC3. Similarly, in PC4, the total variation was 78.08% and the contribution of different traits like days to germination, days to flowering, leaf width, and awn length was recorded as positive contributions toward PC4, and plant height, spike length, node distance, and number of seed per plant was recorded as negative contribution toward PC4.
In order to determine the degree of genetic variety and geographic association among the studied genotypes, the plot generated by the two PCS, PC2 and PC3, showed a total variation of 78.08%. Separation based on PC2 and PC3 revealed that the genotypes were found dispersed throughout each quarter, demonstrating the significant level of genetic variability in the studied genotypes. The genotypes under study were found to be scattered, with the majority of the genotypes grouped in the plot’s center.
Analysis of total seed storage proteins
All the studied genotypes used in morphological characterizations were subjected to SDS-PAGE analysis, where a significant level of variation was recorded. During the present study, 21 polypeptide bands were recorded with molecular weights ranging from 30 to 180 kDa (BLUelf-Prestained Protein ladder) in the 19 studied wheat genotypes. It was found that out of these 21 bands, 18 bands (85.71%) were found to be polymorphic while three bands (14.28%) were found to be monomorphic bands (Figures 5A and C). The electrophorogram of the present studied genotypes is given in Figures 3, 4 and 5C in which considerable genetic diversity can be observed. During the present study, patterns of protein sub-units were studied. Based on the patterns, similarity indices were calculated among all the studied genotypes, and it was found that the similarity ranged from 0.33 (33%) to 1.00 (100%) (Table 4). In the present investigation, the minimum (33%) similarity among the genotypes was Rasol Bnada and Asban, Sandai and Asban, Jannat Banda and Asban, Hayaseri, and Asban, Matta Swat, and Daroo, Matta Swat and Kolal Derai, followed by 0.47 (47%) similarity found among Jannat Banda and Rasool Banda, Jannat Banda, and Sandai. On the other side, the maximum 1.00 (100 %) similarity level was recorded between Dolram and Sadoo, Kolal Derai and Daroo, Sarpato, and Daroo.
Figure 3. Electrophoretic banding pattern of durum wheat genotypes using SDS-PAGE of total seed storage proteins, numbering shows Asban, Dogai, Rasool Banda, Sandai, Kamar kotki, Jannat Banda, Hayaseria, Shaai, Sheikhano, Sadoo, M-Molecular marker.
Figure 4. Electrophoretic banding pattern of durum wheat genotypes using SDS-PAGE of total seed storage proteins, numbering shows Tani Dara, Daroo, Kolal Derai, Rabat, Dol ram, Negotai, Matta Swat, Sarpato, Maklakand, M-Molecular marker.
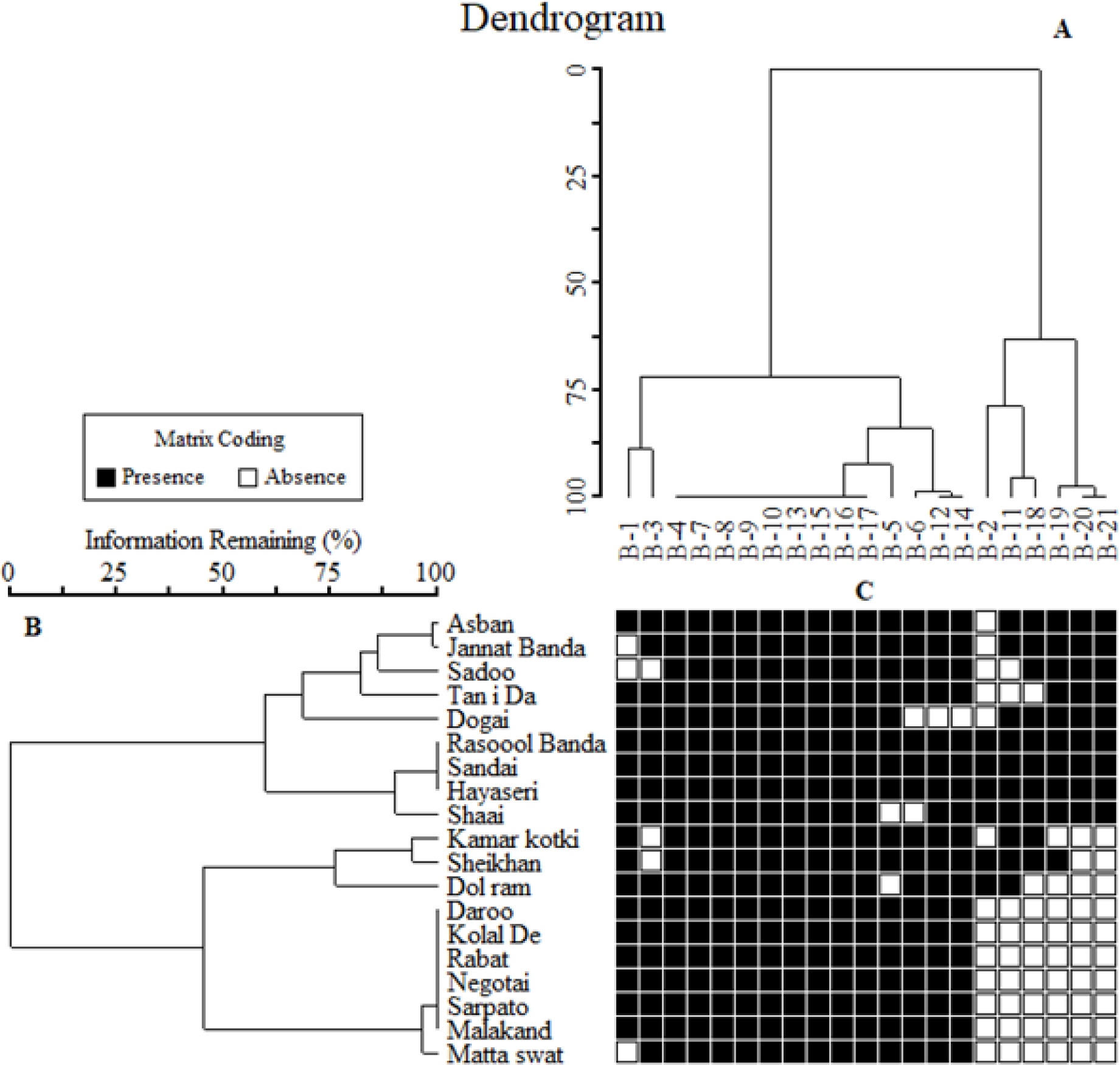
Figure 5. Two-way cluster dendrogram of 19 durum genotypes based on total seed proteins. (A) Shows the diversity of bands, (B) shows the diversity among the genotypes, and (C) shows the presence and absence of bands.
Table 4. Dice coefficient similarity of 19 durum wheat genotypes using SDS-PAGE analysis.
| Dice Coefficients of Similarity | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Genotypes | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 |
| 1 | 1.0 | ||||||||||||||||||
| 2 | 0.57 | 1.00 | |||||||||||||||||
| 3 | 0.33 | 0.66 | 1.00 | ||||||||||||||||
| 4 | 0.33 | 0.66 | 1.00 | 1.00 | |||||||||||||||
| 5 | 0.66 | 0.88 | 0.74 | 0.74 | 1.00 | ||||||||||||||
| 6 | 0.33 | 0.66 | 0.47 | 0.47 | 0.74 | 1.00 | |||||||||||||
| 7 | 0.33 | 0.66 | 1.00 | 1.00 | 0.74 | 0.47 | 1.00 | ||||||||||||
| 8 | 0.57 | 0.66 | 0.47 | 0.47 | 0.88 | 0.66 | 0.47 | 1.00 | |||||||||||
| 9 | 0.66 | 0.88 | 0.57 | 0.57 | 0.47 | 0.74 | 0.57 | 0.74 | 1.00 | ||||||||||
| 10 | 0.57 | 0.81 | 0.66 | 0.66 | 0.74 | 0.47 | 0.66 | 0.81 | 0.74 | 1.00 | |||||||||
| 11 | 0.47 | 0.74 | 0.57 | 0.57 | 0.81 | 0.57 | 0.57 | 0.74 | 0.81 | 0.57 | 1.00 | ||||||||
| 12 | 0.74 | 0.94 | 0.81 | 0.81 | 0.57 | 0.81 | 0.81 | 0.94 | 0.74 | 0.81 | 0.57 | 1.00 | |||||||
| 13 | 0.74 | 0.94 | 0.81 | 0.81 | 0.57 | 0.81 | 0.81 | 0.94 | 0.74 | 0.81 | 0.57 | 1.00 | 1.00 | ||||||
| 14 | 0.74 | 0.94 | 0.81 | 0.81 | 0.57 | 0.81 | 0.81 | 0.94 | 0.74 | 0.81 | 0.57 | 1.00 | 1.00 | 1.00 | |||||
| 15 | 0.81 | 1.00 | 0.74 | 0.74 | 0.66 | 0.88 | 0.74 | 0.74 | 0.66 | 1.00 | 0.81 | 0.57 | 0.57 | 0.57 | 1.00 | ||||
| 16 | 0.74 | 0.94 | 0.81 | 0.81 | 0.57 | 0.81 | 0.81 | 0.94 | 0.74 | 0.81 | 0.57 | 1.00 | 1.00 | 0.00 | 0.57 | 1.00 | |||
| 17 | 0.81 | 1.00 | 0.88 | 0.88 | 0.66 | 0.74 | 0.88 | 1.00 | 0.81 | 0.74 | 0.66 | 0.33 | 0.33 | 0.33 | 0.66 | 0.33 | 1.00 | ||
| 18 | 0.74 | 0.94 | 0.81 | 0.81 | 0.57 | 0.81 | 0.81 | 0.94 | 0.74 | 0.81 | 0.57 | 1.00 | 1.00 | 1.00 | 0.57 | 1.00 | 0.33 | 1.00 | |
| 19 | 0.74 | 0.94 | 0.81 | 0.81 | 0.57 | 0.81 | 0.81 | 0.94 | 0.74 | 0.81 | 0.57 | 1.00 | 1.00 | 1.00 | 0.57 | 1.00 | 0.33 | 1.00 | 1.00 |
Note: 1. Asban, 2. Dogai, 3. Rasoool banda, 4. Sandai, 5. Kamar kotki, 6. Jannat Banda, 7. Hayaseria, 8. Shaai, 9. Sheikhano, 10. Sadoo, 11. Tan i Dara, 12. Daroo, 13. Kolal Derai, 14. Rabat, 15. Dol Ram, 16. Negotai, 17. Matta Swat, 18. Sarpato, 19. Maklakand.
A genetic tree based on a protein binary data matrix divided all the studied wheat genotypes into different clusters based on variation present in their protein profile. A two-way cluster dendrogram was constructed and all the genotypes were divided into two main lineages at a Euclidian distance of 25% and sub-divided into five clusters at a Euclidian distance of 75% where the genotypes Asban and Matta Swat were found to be the most diverse and were placed at the extreme periphery of the dendrogram (Figure 5B). The first sub-cluster consists of four genotypes (Asban Jannat Banda Sadoo and Tan I Da). The sub-cluster two comprised only one genotype Dogai, which shows that it was dissimilar from all the other studied groups and genotypes. The third sub-cluster comprised four genotypes, namely, Rasool Banda, Sandai Hayaseri, and Shaai. The fourth sub-cluster consists of three genotypes (Kamar Kotki, Sheikhan, and Dol Ram). The last sub-cluster five has seven genotypes likely Daroo, Kolal De, Rabat, Negotai, Sarpato, Malakand, and Matta Swat. Such division of all the studied genotypes into the same group and different clusters were due to their similarity and differences level.
Discussion
Morphological characterization is the first step toward the identification and classification of crop germplasm because breeding practices mainly depend upon the magnitude of genetic diversity. Morphological feature is the first step of crop germplasm taxonomy, but the breeding program mainly depends upon genetic characteristics (Ghafoor et al., 2002). That is why it is so important to make proper strategies for the collection and evaluation of germplasm sources, which are locally used in different regions of the world and save them from being vanished. It is the order of the day to investigate the genetic diversity of crop germplasm, explore each aspect of it, and conserve it for use in future breeding programs. The need is to improve the maintenance of the present diversity of different useful crops through characterization and evaluation. At present, crop germplasms are the main sources of genetic diversity, which are used in different breeding programs throughout the globe by plant breeders for the development of new cultivars. In the present study, 19 diverse genotypes were collected from different areas of the Malakand Division of Pakistan and were evaluated for assessing genetic diversity through morphological characterization and total seed storage protein using SDS-PAGE analysis. During the present investigation, a significant level of variation was observed for most of the traits such as days to flowering, days to maturity, plant height, leaf length, node distance, and number of seeds per plant. Early days to germination were recorded for genotype Tangi Dara which germinated in 10 days, followed by Kolal Derai which took 11 days to germinate, while the genotypes Malakand and Dogai were found to be the late germinated genotypes. Days to flowering is one of the most important traits for plant breeders. In the present study, the early flowering genotypes were Tangi Dara and Negotal, which took 107 days to reach the stage of flowering, while the late flowering genotype was Rabat, which took 114 days to flowering. Days to maturity is also one of the important characteristics for plant breeders; early maturity (165) was observed for genotypes Tangi Dara, Daroo, Kolal Derai, and Malakand while the genotypes Dogai, Rasool Banda, Dol Ram, Matta Swat were found to be the genotypes that mature late. Plant height is one of the most important characteristics, ranging from 46.75 to 73.23 cm. In the present study, the minimum plant height (46.8 cm) was observed for genotype Sheikhano while the tallest genotype was Dogai with a plant height of 73.23 cm. Our study is in close agreement with Khan et al. (2021), where the authors studied eight different quantitative traits and found significant variation for most of the studied parameters. Leaves play an important role in plant development and growth, which directly affects the yield of both cereal and fodder plants. The maximum leaf length (24.75 cm) was recorded for Rabat and the lowest leaf length (10 cm) was recorded for Sarpato and Matta Swat had a leaf length of 15 cm. According to Ahmad et al. (2014), variation in leaf and other important parameters in different genotypes at various locations may be also taken to have diversity in the genetic makeup, mineral nutrients, and different environments. Maximum green yield can also be due to larger leaf size which results in the formation of photosynthesis process (Amanullah et al., 2004). For flag leaf area, high heritability and genetic advance estimates have been reported (Saleem et al. 2016). The node distance was recorded with the range from 14.25 to 19.38 and a coefficient of variance of 9.76%. The most important characteristic of a seed-producing plant is its seed yield per plant as seed is the edible part with high nutritive values. A significant level of variation was found for the number of seeds per plant among the studied genotypes with a range of 17.25–29. The minimum number of seeds per plant (29) was recorded for Jannat Banda, followed by Hayaseri having 34 seeds per plant, while the maximum of 46.25 g was recorded for Dol ram followed by Asban and Kamar Kotki, respectively.
Correlation analysis is important for the selection of traits and breeding methods (Ihsan et al., 2021). Plant height was found to be significantly correlated with days to maturity; spike length was significantly correlated with plant height. The present investigation is comparable with that of Khan et al., 2021, where the contributors studied different barley genotypes and found a significant correlation for different morphological traits. The present study was comparable to that of El-Rawy (2020) and Okii, et al., (2014), where the contributor studied different morphological traits and found significant correlations among different parameters. These findings agreed with Feltaous (2019), who found that all wax characteristics examined in various plant parts had a positive correlation with one another. Malik et al., (2014) found a significant positive association between peduncle wax status and leaf blade wax, leaf-sheath wax, and earwax.
In addition, the cluster analysis demonstrated that there was no correlation between genetic diversity and the geographic region because samples collected from the same region were divided into different clusters, whereas genotypes collected from different regions tended to be clustered together and were related. In the present study, all the morphological traits were subjected to cluster analysis and were classified into two main lineages and subsequently subdivided into six sub-clusters. Plants in a given group varied very little from one another but significantly from plants in other groups. These types would be employed in the hybridization program to produce plants with desired characteristics, at least in the region where the study was conducted. PCA are a reliable method for handling germplasm collections. The study’s use of multivariate grouping techniques has applications for both curators of gene banks and breeders. It is possible to choose representative lines from specific groups for a hybridization program with other authorized varieties as well as to remove unidentified lines and genotypes/accessions from the gene bank that are confusing. Genetic diversity has been used to categorize agronomic kinds, and this, combined with biochemical analysis, will make it easier to manage germplasm and create a core collection (Ghafoor et al., 2002). Therefore, it is possible to group genotypes appropriately by using a variety of assessment approaches (Bauer et al., 2007; Kraic et al., 2009). The findings did, however, demonstrate that cluster analysis based on PCA is a more accurate indication of variations across wheat genotypes than cluster analysis. The selection of the most effective genotypes can benefit from an analysis of genetic diversity and the relationships between various features. To increase the grain yield of wide durum wheat cultivars or lines, screening has also been done. However, selection that is only based on grain production is typically not particularly effective. As a result, it is crucial to identify and modify the traits that affect grain yield because doing so improves breeding effectiveness.
SDS-PAGE is one of the most vital tools for the assessment of genetic variation among different crop genotypes (Ghafoor et al., 2005, Zahoor et al., 2015). SDS-PAGE is a biochemical procedure that is employed to ascertain the genetic composition of distinct plants. It has been shown that genotypes exhibit a wide degree of genetic divergence. In the present study, a total of 21 polymorphic bands were identified and a number of significant diversities were recorded based on total seed storage proteins. All the genotypes were sub-classified into different clusters where the genotypes Kamar kotki and Asbanr were found to be the most diverse genotypes and were placed at the extreme periphery of the dendrogram and may be used in the future for breeding practices. Our study is comparable with that of Ihsan et al., (2021) where the collaborators studied different Oat genotypes for total seed storage proteins and recorded a total of 13 polymorphic sub-units. Our results are also comparable with that of Khan et al., (2021) where the authors studied different barely genotypes for total seed storage proteins and observed significant diversity. The genetic diversity estimates based on seed storage protein were the lowest, according to Fufa et al., (2005), because they were the main determinants of end-use quality, a strongly selected characteristic. Considering the foregoing, it is concluded that seed storage protein profiles may serve as useful markers for cultivar identification, variety registration, pedigree analysis, genetic diversity studies, and the classification of adapted cultivars, enhancing the effectiveness of wheat breeding programs for cultivar development.
Conclusions
The morphological characterization is useful to identify the valuable genotypes with desirable quantitative traits for improving genetic diversity in durum quality and therefore suggested that these markers could be useful for the traits and grouping of genotypes on the basis of their origin and performance. Based on the present study, the genotypes Malakand, Dol ram, and Daroo are being recommended to the farmers for future use to increase crop yield. The marker in the proteomics (SDS-PAGE) study is the BLUelf-Prestained Protein ladder. A total of 21 polypeptide bands were recorded with molecular weights ranging from 30 to 180 kDa in the 19 wheat genotypes, with 18 polymorphic bands (85.71%) and 3 monomorphic bands (14.28%). Based on 2-way cluster analysis, all the genotypes were divided into two main lineages where the genotypes Asban and Matta Swat were found to be the most diverse and were placed at the extreme periphery of the dendrogram and may be used in future breeding practices. In the future, the selected genotypes should be examined in salt-rich areas.
Conflict of Interest
The authors declare no conflict of interest.
Acknowledgements
The authors are thankful to the Deanship of Scientific Research (DSR) at King Faisal University for supporting this research work under (Ambitious Researcher Track) with Project no GRANT 5197.
REFERENCES
Aghaee M., Mohammadi R. and Nabovati S. 2010. Agro-morphological characterization of durum wheat accessions using pattern analysis. Australian Journal of Crop Science. 4(7):505–514. 10.3316/informit.536537388993889
Ahmad M., Dar Z.A. and Habib M. 2014. A review on oat (Avena sativa L.) as a dual-purpose crop. Scientific Research and Essays. 9(4):52–59. 10.5897/SRE2014.5820
Awulachew M.T. 2020. Understanding basics of wheat grain and flour quality. Journal of Health and Environmental Research. 6(1):10–26. 10.11648/j.jher.20200601.12
Barcaccia G., 2009. Molecular markers for characterizing and conserving crop plant germplasm. Molecular Techniques in Crop Improvement: 2nd Edition, pp. 231–254. 10.1007/978-90-481-2967-6_10
Bauer I., Drinic S.M., Drinić G. and Micić D.I. 2007. Assessing temporal changes in genetic diversity of maize hybrids using RAPD markers. Cereal Research Communications. 35(4):1563–1571. 10.1556/CRC.35.2007.4.3
El-Rawy M.A. 2020. Assessment of genetic diversity for some Egyptian wheat varieties based on morphological characters and SSR markers. Scientific Journal of Agricultural Sciences. 2(2):144–160. 10.21608/SJAS.2020.37530.1033
Feuillet C., Langridge P. and Waugh R. 2008. Cereal breeding takes a walk on the wild side. Trends in genetics. 24(1): 24–32. 10.1016/j.tig.2007.11.001
Fufa H.U.N.D.E.R.A., Baenziger P.S., Beecher B.S., Dweikat I., Graybosch R.A. and Eskridge K.M. 2005. Comparison of phenotypic and molecular marker-based classifications of hard red winter wheat cultivars. Euphytica. 145: 133–146. 10.1007/s10681-005-0626-3
Ghafoor A., Ahmad Z., Qureshi A.S. and Bashir M. 2002. Genetic relationship in Vigna mungo (L.) Hepper and V. radiata (L.) R. Wilczek based on morphological traits and SDS-PAGE. Euphytica. 123: 367–378. 10.1023/A:1015092502466
Ghasemi N., Mirfakhraii R.G. and Abbasi A. 2019. Assessment of genetic diversity of bread wheat (Triticum aestivum L.) cultivars using microsatellite markers. Journal of Crop Breeding. 11(29): 9–16. 10.29252/jcb.11.29.9
Hassan M.I. 2016. Assessment of genetic diversity in bread wheat genotypes based on heat tolerance and SSR markers. Assiut Journal of Agricultural Sciences. 47(5). 10.21608/AJAS.2016.1987
Ihsan M., Nazir N., Ghafoor A., Khalil A.A.K., Zahoor M., Nisar M., et al. 2021. Genetic diversity in local and exotic Avena sativa L. (Oat) germplasm using multivariate analysis. Agronomy. 11(9): 1713. 10.3390/agronomy11091713
Kesralikar M., Mokate S., Golakiya B.A. and Vakharia D.N. 2017. Genetic diversity analysis among onion genotypes through RAPD, SSR and ISSR markers. International Journal of Current Microbiology and Applied Sciences. 6(6): 1467–1474. 10.20546/ijcmas.2017.606.172
Khaled A.G.A., Motawea M.H. and Said A.A. 2015. Identification of ISSR and RAPD markers linked to yield traits in bread wheat under normal and drought conditions. Journal of Genetic Engineering and Biotechnology. 13(2): 243–252. 10.1016/j.jgeb.2015.05.001
Khan A., Ihsan M., Nisar M., Hazrat A., Ali M., Ul-Haq R., et al. 2021. Evaluation of Genetic Diversity in Barley Landraces through Agro-Morphological and Biochemical Characterization. Sarhad Journal of Agriculture. 37. 10.17582/journal.sja/2021/37.3.984.992
Khan J., Khurshid S., Sarwar A., Aziz T., Naveed M., Ali U., et al. 2022. Enhancing Bread Quality and Shelf Life via Glucose Oxidase Immobilized on Zinc Oxide Nanoparticles—A Sustainable Approach towards Food Safety. Sustainability. 14(21): 14255. 10.3390/su142114255
Khan M.K., Pandey A., Thomas G., Akkaya M.S., Kayis S.A., Ozsensoy Y., et al. 2015. Genetic diversity and population structure of wheat in India and Turkey. AoB Plants. 7: 83. 10.1093/aobpla/plv083
Khodadadi M., Fotokian M.H. and Miransari M., 2011. Genetic diversity of wheat (Triticum aestivum L.) genotypes based on cluster and principal component analyses for breeding strategies. Australian Journal of Crop Science. 5(1): 17–24.
Kraic F.I.L.I.P., Mocák J., Roháčik T.I.B.O.R. and Sokolovičová J.A.N.A. 2009. Chemometric characterization and classification of new wheat genotypes. Nova Biotechnol. 9:101–106.
Malik R., Sharma H., Sharma I., Kundu S., Verma A., Sheoran S., Kumar R. and Chatrath R. 2014. Genetic diversity of agro--morphological characters in Indian wheat varieties using GT biplot. Australian Journal of Crop Science. 8(9): 1266–1271.
Niaz S., Ahmad H., Khan I.A., Shah A.H. and Rahman I. 2014. Molecular and agronomic characterization of durum wheat. Intl. J. Biosci. 4: 270–275. 10.12692/ijb/4.7.270-275
Nisar M., Ghafoor A., Khan M.R. and Subhan M. 2009. Genetic similarity of Pakistani pea (Pisum sativum L.) germplasm with world collection using cluster analysis and jaccard’s similarity index. Journal of the Chemical Society of Pakistan. 31(1): 138–144.
Okii D., Tukamuhabwa P., Odong T., Namayanja A., Mukabaranga J., Paparu P. and Gepts P., 2014. Morphological diversity of tropical common bean germplasm. African Crop Science Journal. 22(1): 59–68.
Saleem B., Khan A.S., Shahzad M.T. and Ijaz F. 2016. Estimation of heritability and genetic advance for various metric traits in seven F2 populations of bread wheat (Triticum aestivum L.). Journal of Agricultural Sciences. 61(1): 1–9. 10.2298/JAS1601001S
Salem K.F., Röder M.S. and Börner A. 2015. Assessing genetic diversity of Egyptian hexaploid wheat (Triticum aestivum L.) using microsatellite markers. Genetic resources and crop evolution. 62: 377–385. 10.1007/s10722-014-0159-5
Shuaib M., Zeb A., Ali Z., Ali W., Ahmad T. and Khan I. 2007. Characterization of wheat varieties by seed storage protein electrophoresis. African Journal of Biotechnology. 6(5).
Siahbidi M.M.P., Aboughadareh A.P., Tahmasebi G.R., Teymoori M. and Jasemi M. 2013. Evaluation of genetic diversity and interrelationships of agro-morphological characters in durum wheat (Triticum durum Desf.) lines using multivariate analysis. International Journal of Agriculture. 3(1): 184.
Varzakas T., Kozub N. and Xynias I.N. 2014. Quality determination of wheat: genetic determination, biochemical markers, seed storage proteins–bread and durum wheat germplasm. Journal of the Science of Food and Agriculture. 94(14): 2819–2829. 10.1002/jsfa.6601
Zahoor M., Nisar M. and Islam N. 2015. Genetic variations of Robinia pseudoacacia plant using SDS-PAGE. Pak. J. Bot. 46(6): 2335–2338.
Zarkti H., Ouabbou H., Hilali A. and Udupa S.M. 2010. Detection of genetic diversity in Moroccan durum wheat accessions using agro-morphological traits and microsatellite markers. African Journal of Agricultural Research. 5(14): 1837–1844. 10.5897/AJAR09.249
Zhang W. and Cai X. 2019. Alien introgression and breeding of synthetic wheat. In Advances in breeding techniques for cereal crops (pp. 3–54). Burleigh dodds science publishing.
Zubair M., Ajmal S.U., Anwar M. and Haqqani A.M. 2007. Multivariate analysis for quantitative traits in mungbean [Vigna radiata (L.) Wilczek]. Pakistan Journal of Botany. 39(1): 103.
Supplementary
Table S1. Passport information of studied genotypes (Landraces) used in the present study.
| S. No | Name | Scientific name | Origin |
|---|---|---|---|
| 1 | Asban | Triticum durum | KPK Pakistan |
| 2 | Dogai | Triticum durum | KPK Pakistan |
| 3 | Rasoool bada | Triticum durum | KPK Pakistan |
| 4 | Sandai | Triticum durum | KPK Pakistan |
| 5 | Kamar kotki | Triticum durum | KPK Pakistan |
| 6 | Jannat Banda | Triticum durum | KPK Pakistan |
| 7 | Sheikhano | Triticum durum | KPK Pakistan |
| 8 | Daroo | Triticum durum | KPK Pakistan |
| 9 | Kolal Derai | Triticum durum | KPK Pakistan |
| 10 | Hayaseria | Triticum durum | KPK Pakistan |
| 11 | Tan i Dara | Triticum durum | KPK Pakistan |
| 12 | Sadoo | Triticum durum | KPK Pakistan |
| 13 | Sha ai | Triticum durum | KPK Pakistan |
| 14 | Rabat | Triticum durum | KPK Pakistan |
| 15 | Dol ram | Triticum durum | KPK Pakistan |
| 16 | Negotal | Triticum durum | KPK Pakistan |
| 17 | Matta swat | Triticum durum | KPK Pakistan |
| 18 | Sarpato | Triticum durum | KPK Pakistan |
| 19 | Malakand | Triticum durum | KPK Pakistan |