
Download
PAPER
Marker-assisted selection of dairy cows for β-casein gene A2 variant
Carla Sebastiani1†, Chiara Arcangeli1†, Martina Torricelli1*, Marcella Ciullo1, Nicoletta D’avino1, Giulia Cinti2*, Stefano Fisichella1, Massimo Biagetti1
1Istituto Zooprofilattico Sperimentale dell’Umbria e delle Marche-Togo Rosati (IZSUM), Perugia, Italy
2R&D Cooperlat, Società Cooperativa Agricola, Jesi, Ancona, Italy
Abstract
Many studies highlighted potential associations of β-casein A1 with specific human diseases and a minor digestibility of milk, due to the bioactive peptide β-casomorphin 7 (BCM-7) release during digestion. Conversely, the ancestral β-casein A2 variant seems to be a favorable trait because it is not associated with BMC-7 release. The aim of this work was to evaluate frequencies of β-casein variants in offspring of previously genotyped cows inseminated with A2 homozygous semen. The frequency of the A2/A2 animals has almost doubled from 37 to 69%. These are encouraging results with the perspective of reaching the goal of producing A2 milk.
Key words: β-casein, bovine, marker-assisted selection (MAS), milk, polymorphisms, variants
*Corresponding Author: Martina Torricelli, Istituto Zooprofilattico Sperimentale dell’Umbria e delle Marche-Togo Rosati (IZSUM), Via Salvemini 1, 06126 Perugia, Italy. Email: [email protected]; Giulia Cinti, R&D Cooperlat, Società Cooperativa Agricola, via Piandelmedico 74, 60035 Jesi (Ancona), Italy. Email: [email protected]
†These authors contributed equally.
Received: 25 January 2022; Accepted: 25 February 2022; Published: 2 April 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Marker-Assisted Selection (MAS) is a methodology that allows for the selection of important genetic animal traits in the population of interest by exploiting the genetic information at specific markers. Advancement in genomics made easier the identification of markers which could be ultimately utilized in MAS. In particular, the Genome Wide Association Study (GWAS) approach, initially used in human genetics research to associate genetic variations with particular diseases, is of special interest (Raina et al., 2020). The method is based on scanning the genome of many different individuals for genetic markers that can be used to predict the presence of a disease in the population under study. More recently, GWAS has been applied to the field of domestic animal breeding and genetics, and many genetic markers affecting important economical traits have been described (Du et al., 2021; Sharma et al., 2015).
Cattle breeding programs, consisting of marker-assisted selection (MAS), has been applied to the selection of dairy cows for the presence of β-casein A2 variant, known to confer better digestibility to milk (Duarte-Vazquez et al., 2017; Kaminski et al., 2007; Park et al., 2021).
Milk and dairy products are among the main components of the food tradition of many countries and they play a fundamental role in human health due to their valuable nutritional properties. In spite of various sources of milk being available on the market, bovine milk represents the most consumed variety of milk worldwide (Faye and Konuspayeva, 2012). It contains essential nutrients such as proteins with high biological value, lipids, carbohydrates (mainly lactose), minerals (calcium, phosphorus, zinc, and magnesium), and vitamins (i.e., B2, B12, D, and A) (Jenness et al., 1979; Muehlhoff et al., 2013). Regarding milk protein fraction, it is composed by soluble proteins, also known as whey proteins, and by insoluble proteins, that are caseins, which represent about 80% of the total bovine milk proteins. These are subdivided into four groups, αs1, αs2, β, and k, encoded respectively by the CSN1S1, CSN1S2, CSN2, and CSN3 genes, all located on chromosome 6 (Rijnkels, 2002). In spite of high milk consumption all over the world, some people experience digestive disorders following the intake of milk and dairy products, because of lactose malabsorption or digestive difficulties due to other dairy components such as β-casein, which represents about 36% of milk protein content (Milan et al., 2020). Specifically, some studies have shown a correlation between human health and some β-casein variants (Kay et al., 2021; Thiruvengadam et al., 2021). Indeed, Bos taurus CSN2 gene harbors many nucleotide substitutions leading to the formation of 12 protein variants (A1, A2, A3, B, C, D, E, F, G, H1, H2, and I), seven of which (A1, A2, A3, B, C, I, and E) have been identified mainly in European cattle breeds (Barroso et al., 1999; Daniloski et al., 2021; Hohmann et al., 2021; Massella et al., 2017) (Table 1).
Table 1. Differences in the amino acid sequence of β-casein variants.
| β-casein Variant |
Amino Acid Position | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| 36 | 37 | 67 | 72 | 88 | 93 | 106 | 122 | 138 | |
| A2* | Glu (E) | Glu (E) | Pro (P) | Gln (Q) | Leu (L) | Met (M) | His (H) | Ser (S) | Pro (P) |
| A1* | His (H) | ||||||||
| A3* | Gln (Q) | ||||||||
| B* | His (H) | Arg (R) | |||||||
| C* | Lys (K) | His (H) | |||||||
| E* | Lys (K) | ||||||||
| I* | Leu (L) | ||||||||
| D | |||||||||
| F | His (H) | Leu (L) | |||||||
| G | His (H) | Leu (L) | |||||||
| H1 | Ile (I) | ||||||||
| H2 | Glu (E) | Leu (L) | Glu (E) | ||||||
In bold: amino acid variations with respect to the A2 ancestral variant.
Arg: arginine; Gln: glutamine; Glu: glutamic acid; His: histidine; Ile: isoleucine; Leu: leucine; Lys: lysine; Met: methionine; Pro: proline; Ser: serine;
*Allele variants detected in European cattle breeds.
Among the dairy cattle breeds, A1 and the ancestral A2 variants are the most common while B and I variants are generally less frequent with some variability depending on the breed (Farrel et al., 2004). Other variants, such as A3 and C, are rarely found (Farrel et al., 2004; Massella et al., 2017) and others are related to specific breeds or geographic areas, as it happens for the E and F variants that have been found with a very low frequency, only in the Italian Piedmontese breed and in animals reared in the Emilia-Romagna region (northern Italy), respectively (Massella et al., 2017; Voglino et al., 1972).
In the last two decades, much attention has been paid to the A1 and A2 β-casein content of milk, due to their suggested role in human health. A1 and A2 variants differ from each other at the gene level for a point mutation, which causes an amino acid change. Particularly, at position 67 of the protein chain, a histidine in the A1 variant is replaced by a proline in the A2 variant. Because of this difference in the protein sequence, the β-casein variants A1 and A2 are differently processed during digestion. Actually, digestive enzymes perform a proteolytic cleavage at position 67 of the β-casein chain only when a histidine is present generating a seven amino acids peptide named β-casomorphin 7 (BCM-7), while cleavage is prevented by the presence of a proline at the same position. Other variants are characterized by a proline at position 67 (A3, E, D, I, and H) and could exhibit the same behavior of the A2 variant as well as other variants, with a histidine at the same position (B, C, F, and G), and could behave as the A1 variant resulting in the formation of the β-casomorphin 7 (BCM-7) (Bodnar et al., 2018).
BMC-7 is a bioactive peptide with strong opioid activity and an oxidant effect (EFSA Scientific Report 2009; Kay et al., 2021) and its release has been related to the alteration of the physiology of different organ systems. In particular, several studies demonstrated the correlation with the onset of various human pathological conditions, such as heart disease, sudden infant death syndrome, milk intolerance, and also with the aggravation of symptoms associated with schizophrenia, autism, and type 1 diabetes (Caroli et al., 2009; Cieslinska et al., 2015; Kaminski et al., 2007; Kay et al., 2021; McLachlan, 2001; Pal et al., 2015; Reichelt et al., 2012). In more detail, it has been reported in the literature that BMC-7, binding to µ receptor in the gastrointestinal (GI) tract, may alter gut microbiota, can increase inflammatory response, and may induce mucin production, thus triggering lactose intolerance-like symptoms. Moreover, BMC-7 may promote oxidative stress, may deregulate insulin metabolism, and can modulate DNA methylation reactions affecting neurodevelopment (Kay et al., 2021). However, the European Food Safety Authority (EFSA) in 2009 carried out a meta-analysis of data present in the literature, releasing a scientific report that supports the absence of a cause–effect relationship between the consumption of milk containing the A1 variant and the etiology of the aforementioned diseases, so further studies are necessary in this field (EFSA Scientific Report, 2009). Therefore, the discussion on the adverse human health effects of the β-casein variant A1 remains still open also because other studies demonstrated that milk obtained from A2 homozygous cows seems to be more digestible than milk containing β-casein A1 (Deth et al., 2016; He et al., 2017; Ramakrishnan et al., 2020). This effect could be traced back to the increase in the rate of gastrointestinal transit of A2 milk and to the lack of the pro-inflammatory effect instead associated with A1 milk consumption (Brooke-Taylor et al., 2017; Kay et al., 2021). In addition, A2 milk consumption increases the natural production of glutathione (GSH), which is reported to be a key antioxidant, widely recognized for its association with many health benefits. The consumption of A2 milk induces a twofold increase of blood GSH levels compared to the levels derived from conventional milk intake (Deth et al., 2016). Interestingly, human breast milk, which is recommended by the World Health Organization (WHO) as the exclusive food for newborn feeding, is characterized by the presence of a β-casein protein carrying a proline residue in position 67 which is therefore very similar to the bovine A2 variant (Kay et al., 2021).
As a consequence, in many countries, including Australia, the United Kingdom, the United States, New Zealand, the Netherlands, China and more recently also Italy, A2 cow’s milk has been made commercially available, and it is widely recommended for people who suffer from milk-intolerance and for newborns who need formulas more soft to their digestive system (Brooke-Taylor et al., 2017).
Considering this scenario, this study focused on the planning and execution of a breeding program based on the MAS selection of the β-casein A2 variant, in farms located in central Italy providing milk for an important drinking milk producing plant. A2 heterozygous and homozygous cows, previously genotyped (Sebastiani et al., 2020), were artificially inseminated with semen from bulls homozygous for the A2 variant, and their offspring have been likewise analyzed in order to identify A2/A2 animals for A2 milk production.
Materials and methods
Sampling
A total of 1452 Italian Holstein Friesian cows, reared in farms located in central Italy and previously genotyped (Sebastiani et al., 2020), were subjected to artificial insemination with semen from A2/A2 selected bulls (Co.S.A.P.A.M. Soc. Coop., Lodi, Italy; ABS Italia Srl, Cremona, Italy; INSEME Spa, Modena, Italy). Among these, 640 were A2 homozygous and 812 were A2 carriers (A1/A2, A2/B, A2/I) animals.
From the pregnant cows, 534 heifers were born. From these animals, whole blood samples were collected in tubes containing ethylenediaminetetra-acetic acid (EDTA) as an anticoagulant and stored at −20 °C until genetic analysis. Samples were taken in a single withdrawal, simultaneously with the mandatory periodic tests required by Italian National Health Programs and during breeders’ voluntary health controls.
DNA extraction and sequencing
Genomic DNA was extracted using High Pure PCR Template Preparation Kit (Roche Life Science, Mannheim, Germany) according to the manufacturer’s instructions. PCR reactions of both exons 6 and 7 were performed as previously described (Sebastiani et al., 2020). Amplicons were analyzed through 2% agarose gel electrophoresis containing Midori Green Advanced DNA Stain (Nippon Genetics Europe GmbH, Düren, Germany). PCR products were purified with QIAquick® PCR Purification Kit (Qiagen, Hilden, Germany) and sequenced in both directions using BrilliantDyeTM Terminator Cycle Sequencing Kit v3.1 (NimaGen BV, Nijmegen, Netherlands) according to the manufacturer’s instructions.
Sequencing reactions were analyzed in a 3500 Genetic Analyzer (Applied Biosystems; Thermo Fisher Scientific Inc.). The obtained nucleotide sequences were aligned to the bovine β-casein gene (Accession number X14711.1) using the ClustalW tool of the BioEdit v7.2.5 software (Hall, 1999). Electropherograms were analyzed at each investigated mutation point to identify peaks in heterozygosity. In particular, polymorphisms at positions 36 and 37 of exon 6 and at positions 67, 72, 88, 93, 106, 122, and 138 of exon 7 were analyzed to discriminate the different β-casein variants.
Statistical analysis
Allele and genotype frequencies were directly calculated dividing the number of copies of each allele and genotype by the total alleles and by the total individuals, respectively. Furthermore, the Hardy–Weinberg (HW) equilibrium was verified using Chi-square test (P < 0.05) by the R Studio software (R Core Team, 2020).
Results
In this study, 1452 already genotyped dams (Sebastiani et al., 2020), at least carrier of the A2 allele (A2 heterozygous and homozygous animals), were artificially inseminated using commercial semen from A2 homozygous bulls in order to increase A2 variant and A2/A2 genotype frequencies in the female progeny.
From inseminated cows that got pregnant, 534 heifers were born. Among these, 238 derived from A2 homozygous parents were analyzed to confirm the A2/A2 genotype as they have to be used for the production of A2 certified milk. The remaining 296 heifers, born from A2 heterozygous dams, were analyzed in order to define their genotype and separate the A2 homozygous ones in the herds with the same purpose.
Here, we report the results of the analysis carried out on this offspring in terms of allele and genotype frequencies of the different CSN2 gene variants. No deviation of HW equilibrium was observed at the considered polymorphic sites.
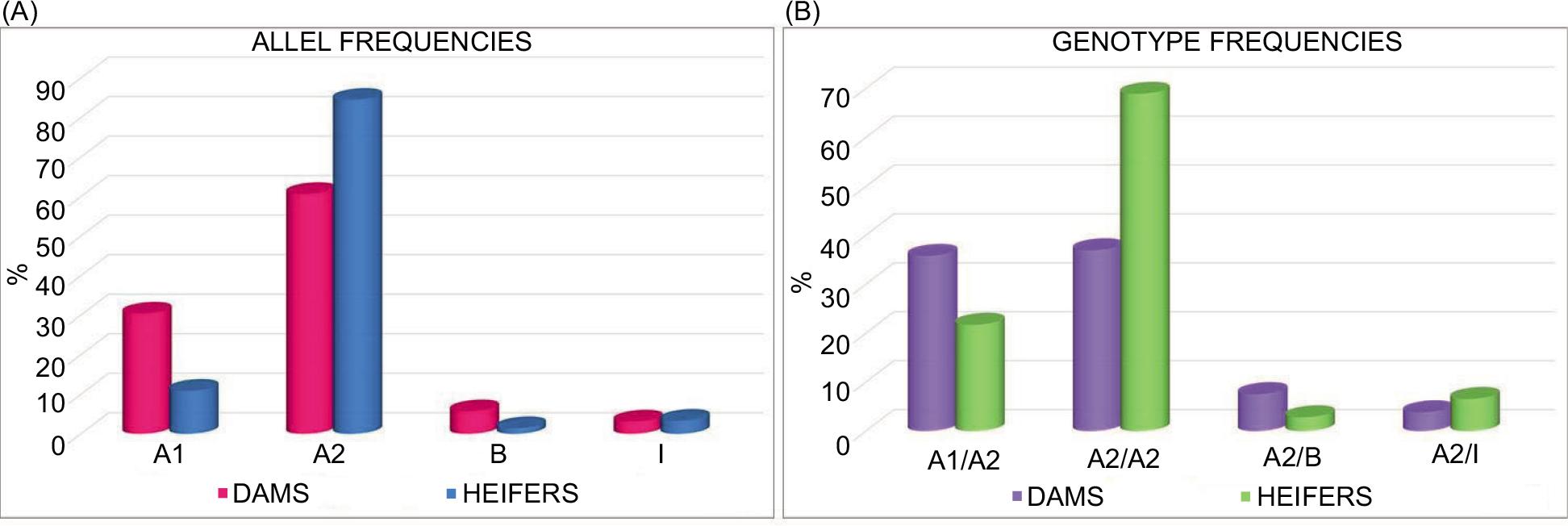
In the female offspring population obtained by the artificial insemination of A2 heterozygous and homozygous cows, sequencing analysis of the CSN2 gene PCR products highlighted the presence of four β-casein variants (A1, A2, B, and I) and four genotypes, (A2/A2, A1/A2, A2/B, A2/I) as shown in Table 2. After the application of the MAS on the herds participating in the project, the frequencies of A2 and I alleles, both characterized by a proline in position 67, increased from 60.65 to 84.46% and from 3.10 to 3.28%, respectively, in the progeny compared to dams. At the same time, the frequencies of the unfavorable A1 and B alleles decreased from 30.39 to 10.86% and from 5.68 to 1.40%, respectively. A3 and C alleles and their related genotypes, that were present with low frequencies in the population of dams, were not further found in the offspring. Regarding genotypes, the most interesting result concerned the about twofold increase of A2/A2 frequency from 36.96 to 68.91%. A similarly intriguing result, derived from the MAS application, was the consistent reduction of the frequency of A1/A2 animals, from 35.79 to 21.72%. The frequencies of the other A2 heterozygous genotypes found in the progeny, that is A2/I and A2/B, varied from 7.55 to 2.81% and from 3.83 to 6.55% (Table 2, Figure 1). Interestingly, since the I variant should behave in the same manner as A2 variant in term of BMC-7 formation, A2/I animals could at least be used for the production of a more digestible milk, even though they could not contribute to the marketing of a certified A2 milk.
Table 2. Allele and genotype frequencies (%) in the examined animals, before (dams) and after (heifers) MAS selection.
| Allele | Allele frequency (%) | Genotype | Genotype frequency (%) | ||
|---|---|---|---|---|---|
| Dams | Heifers | Dams | Heifers | ||
| A2 | 60.65 | 84.46 | A2/A2 | 36.96 | 68.91 |
| A1 | 30.39 | 10.86 | A1/A2 | 35.79 | 21.72 |
| B | 5.68 | 1.40 | A1/A1 | 9.88 | / |
| I | 3.10 | 3.28 | A2/B | 7.55 | 2.81 |
| A3 | 0.15 | / | A2/I | 3.83 | 6.55 |
| C | 0.03 | / | A1/B | 3.07 | / |
| A1/I | 2.03 | / | |||
| B/I | 0.25 | / | |||
| B/B | 0.18 | / | |||
| A2/A3 | 0.12 | / | |||
| A3/B | 0.12 | / | |||
| A1/A3 | 0.06 | / | |||
| A1/C | 0.06 | / | |||
Figure 1. Graphics of the allele (A) and genotype (B) frequencies variation (%) in the examined dams and heifers after MAS selection.
In conclusion, the first generation from the genotyped dams produced an extra number of 368 A2/A2 cows in addition to the A2 homozygous cows in reproductive age still present in the herds. So, both groups of animals may be used for reproduction and for the related A2 drinking milk production.
Discussion
The data presented here were derived from a larger research project whose purpose was primarily to evaluate the frequencies of the β-casein CSN2 gene alleles in Italian Holstein Friesian dairy cattle reared in farms supplying milk to an important milk plant in central Italy.
In this study, we wanted to evaluate the increase of the A2 allele and A2/A2 genotype frequencies, in the female progeny of dams, previously genotyped and fertilized with A2/A2 semen in a selection program based on MAS. The final goal was the production of A2 cow’s milk, due to its supposed association with health benefits for humans.
In fact, the presence in milk of the A2 isoform of β-casein is increasingly considered a desirable characteristic, because it confers greater digestibility to milk. This could allow milk intake even by people who suffer from lactose intolerance-like symptoms despite not being really lactose intolerant (Park et al., 2021).
The Italian Holstein Friesian breed is not among those breeds characterized by the highest frequency of the A2/A2 genotype, but it has a sufficiently high frequency to allow an effective genetic selection of this trait (Canavesi, 2016).
Production of A2 milk can be accomplished following a MAS scheme, which is based on selecting animal carriers of specific polymorphisms that characterize the A2 variant.
During the course of a MAS-based selection plan, aimed at obtaining a consistent number of A2 homozygous cows in the herd, milk coming from A2/A2 animals can be collected separately and directed to A2 milk commercialization. This approach requires organizing, logistical, and management efforts of cow sheds and milking barns, which however is rewarded by an economic gain in the sale of a type of milk with beneficial properties for human health.
In the last years, the commercialization of A2 milk has conquered large market portions in many non-European countries, while in Europe and in Italy it still remains a niche product (Brooke-Taylor et al., 2017). Actually, few medium/large Italian dairy industries have taken this route commercializing certified A2 milk, whose distribution is slowly spreading, starting to generate the interest of consumers.
The data reported here are very encouraging because they confirm the expected genetic improvement in the farms analyzed in this survey. The use in the future of certified A2/A2 sexed semen could help to accelerate the increase in the number of female animals to be used for A2 milk production.
Conclusions
In recent decades, advances in assisted reproductive technologies, animal molecular genetics, and statistical analysis applied to animal genetic improvement helped to maximize the genetic gain in livestock breeding worldwide.
In the past, cows produced milk whose β-casein protein was represented only by A2 isoform, considered to be ancestral, but over time changes in the genetic heritage led to the occurrence of other variants (Farrel et al., 2004). Cows have thus acquired the ability to produce milk with different β-casein isoforms, in particular A1 and A2.
Nowadays, β-casein A2 milk can be considered “a return to the origins” because it comes from selected cows that produce only the ancestral β-casein A2 protein. It has to be noted that in addition to the indexes evaluated for the selection of the best breeding bulls regarding traits related to morphology, health, and productivity, information about milk quality, such as β-casein genotype, has also been included.
In conclusion, the valorization of this genetic trait and production of A2 milk could be advantageous for the whole drinking milk chain, from producers to consumers.
Author Contributions
S.F., G.C., and M.B. conceptualized the study; C.S. and M.B. formulated the study; C.A. and C.S. did the formal analysis; C.A., M.T., M.C., and C.S. were in charge of investigation; G.C., S.F., and N.D. arranged the resources; C.A. and C.S. prepared the original draft; M.B., C.S., M.T., C.A, and G.C. reviewed and edited the manuscript; M.B. and C.S. supervised the study; M.B., G.C., and S.F. were in charge of project administration; and S.F. and G.C acquired funding.
Disclosure Statement
All authors report no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Funding
This research was funded by Progetto Stalla 20.20—ID Progetto n. 21999—Misura 16.2-PSR Marche 2014-2020–Filiera Agro-Alimentare n. 497—Coordinator: Cooperlat Soc. Coop. Agr.
REFERENCES
Barroso, A., Dunner, S. and Cañón, J., 1999. Technical note: use of PCR-single-strand conformation polymorphism analysis for detection of bovine β-casein variants A1, A2, A3, B. J. Anim. Sci. 77(10): 2629–2632. 10.2527/1999.77102629x
Bodnár, Á., Hajzser, A., Egerszegi, I., Póti, P., Kuchtík, J. and Pajor, F., 2018. A2 milk and its importance in dairy production and global market. Anim. Welf. 14(1): 1–7. 10.17205/SZIE.AWETH.2018.1.001
Brooke-Taylor, S., Dwyer, K., Woodford, K. and Kost, N., 2017. Systematic review of the gastrointestinal effects of A1 compared with A2 β-Casein. Adv. Nutr. 15(5): 739–748. 10.3945/an.116.013953
Canavesi, F., 2016. Selezionare per produrre latte A2. Professione allevatore. 16: 52–54.
Caroli, A.M., Chessa, S. and Erhardt, G.J., 2009. Invited review: milk protein polymorphism in cattle: effect on animal breeding and human nutrition. J. Dairy Sci. 92(11): 5335–5352. 10.3168/jds.2009-2461
Cieslinska, A., Sienkiewicz-Szłapka, E., Wasilewska, J., Fiedorowicz, E., Chwała, B., Moszy’nska-Dumara, M. and Kostyra, E., 2015. Influence of candidate polymorphisms on the dipeptidyl peptidase IV and μ-opioid receptor genes expression in aspect of the β-casomorphin-7 modulation functions in autism. Peptides 65: 6–11. 10.1016/j.peptides.2014.11.012
Daniloski, D., Cunha, N.M.D., McCarthy, N.A., O’Callaghan, T.F., McParland, S. and Vasiljevic, T., 2021. Health-related outcomes of genetic polymorphism of bovine β-casein variants: a systematic review of randomized controlled trials. Trends Food Sci. Technol. 111: 233–248. 10.1016/j.tifs.2021.02.073
Deth, R., Andrew Clarke, A., Jiayi, N.J. and Trivedi, M., 2016. Clinical evaluation of glutathione concentrations after consumption of milk containing different subtypes of β-casein: results from a randomized, cross-over clinical trial. Nutr. J. 15(1): 82–87. 10.1186/s12937-016-0201-x
Du, L., Duan, X., An, B., Chang, T., Liang, M., Xu, L., Zhang, L., Li, J., Guangxin, E. and Gao, H., 2021. Genome-wide association study based on random regression model reveals candidate genes associated with longitudinal data in Chinese Simmental Beef Cattle. Animals. 11(9): 2524. 10.3390/ani11092524
Duarte-Vázquez, M.Á., García-Ugalde, C., Villegas-Gutiérrez, L.M., García-Almendárez, B.E. and Rosado, J.L., 2017. Production of cow’s milk free from beta-casein A1 and its application in the manufacturing of specialized foods for early infant nutrition. Foods. 6(7): 50. 10.3390/foods6070050
Farrel, H.M., Jimenez-Flores, R., Bleck, G.T., Brown, E.M., Butler, J.E., Creamer, L.K. and Swaisgood, H.E., 2004. Nomenclature of the proteins of cows’ milk—sixth revision. J. Dairy Sci. 87(6): 1641–1674. 10.3168/jds.S0022-0302(04)73319-6
Faye, B. and Konuspayeva, G., 2012. The sustainability challenge to the dairy sector–the growing importance of non-cattle milk production worldwide. Int Dairy J. 24(2): 50–56. 10.1016/j.idairyj.2011.12.011
Hall, T.A., 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41: 95–98.
He, M., Sun, J., Jiang, Z.Q. and Yang, Y.X., 2017. Effects of cow’s milk beta-casein variants on symptoms of milk intolerance in Chinese adults: a multicenter, randomized controlled study. Nutr. J. 16(1): 72. 10.1186/s12937-017-0275-0
Hohmann, L.G., Weimann, C., Scheper, C., Erhardt, G. and König, S., 2021. Genetic diversity and population structure in divergent German cattle selection lines on the basis of milk protein polymorphisms. Arch. Anim. Breed. 64(1): 91–102. 10.5194/aab-64-91-2021
Jenness, R., 1979. Comparative aspects of milk protein. J. Dairy Res. 46(2): 197–210. 10.1017/s0022029900017040
Kamiński, S., Cieślińska, A. and Kostyra, E., 2007. Polymorphism of bovine β-casein and its potential effect on human health. J. Appl. Genet. 48(3): 189–198. 10.1007/BF03195213
Kay, S.-I.S, Delgado, S., Mittal, J., Eshraghi, R.S., Mittal, R. and Eshraghi, A.A., 2021. Beneficial effects of milk having A2 β-casein protein: myth or reality? J. Nutr. 151(5): 1062–1072. 10.1093/jn/nxaa454
Massella, E., Piva, S., Giacometti, F., Liuzzo, G., Zambrini, A.V. and Serraino, A., 2017. Evaluations of bovine β-casein polymorphism in two dairy farms located in northern Italy. Ital. J. Food Safety. 6(3): 6904. 10.4081/ijfs.2017.6904
McLachlan, C.N., 2001. β-casein A1, ischaemic heart disease mortality and other illnesses. Med. Hypotheses 56(2): 262–272. 10.1054/mehy.2000.1265
Milan, A.M., Shrestha, A., Karlström, H.J., Martinsson, J.A., Nilsson, N.J., Perry, J.K., Day, L. and Barnett, M.P.G. and Cameron-Smith, D., 2020. Comparison of the impact of bovine milk β-casein variants on digestive comfort in females self-reporting dairy intolerance: a randomized controlled trial. Am. J. Clin. Nutr. 111(1): 149–160. 10.1093/ajcn/nqz279
Muehlhoff, E., Bennett, A. and McMahon, D., 2013. Milk and dairy product in human nutrition. FAO, Rome.
Pal, S., Woodford, K., Kukuljan, S. and Ho, S., 2015. Milk intolerance, β-casein and lactose. Nutrients 7(9): 285–297. 10.3390/nu7095339
Park, Y.W. and Haenlein, G.F.W., 2021. A2 bovine milk and caprine milk as a means of remedy for milk protein allergy. Dairy. 2(2): 191–201. 10.3390/dairy2020017
R Core Team, 2020. R: a language and environment for statistical computing. R Foundation for Statistical Computing, R Core Team, Vienna, Austria.
Raina, V.S., Kour, A., Chakravarty, A.K. and Vohra, V., 2020. Markerassisted selection visàvis bull fertility: coming full circle—a review. Mol. Biol. Rep. 47(11): 9123–9133. 10.1007/s11033-020-05919-0
Ramakrishnan, M., Eaton, T.K., Sermet, O.M. and Savaiano, D.A., 2020. Milk containing A2-casein only, as a single meal, causes fewer symptoms of lactose intolerance than milk containing A1 and A2-caseins in subjects with lactose maldigestion and intolerance: a randomized, double-blind, crossover trial. Nutrients. 12(12): 3855. 10.3390/nu12123855
Reichelt, K.L., Tveiten, D., Knivsberg, A.M. and Brønstad, G., 2012. Peptides’ role in autism with emphasis on exorphins. Microb. Ecol. Health Dis. 23: 18958. 10.3402/mehd.v23i0.18958
Rijnkels, M., 2002. Multispecies comparison of the casein gene loci and evolution of casein gene family. J Mammary Gland Biol. Neoplasia. 7(3): 327–345. 10.1023/A:1022808918013
Scientific Report of EFSA prepared by a DATEX Working Group on the potential health impact of β-casomorphins and related peptides. 2009. EFSA Scientific Report. 231: 1–107. 10.2903/j.efsa.2009.231r
Sebastiani, C., Arcangeli, C., Ciullo, M., Torricelli, M., Cinti, G., Fisichella, S. and Biagetti, M., 2020. Frequencies evaluation of β-casein gene polymorphisms in dairy cows reared in central Italy. Animals. 10(2): 252. 10.3390/ani10020252
Sharma, A., Lee, J.S., Dang, C.G., Sudrajad, P., Kim, H.C., Yeon, S.H., Kang, H.S. and Lee, S.H., 2015. Stories and challenges of genome wide association studies in livestock—a review. Asian Australas. J. Anim. Sci. 28(10): 1371–1379. 10.5713/ajas.14.0715
Thiruvengadam, M., Venkidasamy, B., Thirupathi, P., Chung, I.M. and Subramanian, U., 2021. β-Casomorphin: a complete health perspective. Food Chem. 337: 127765. 10.1016/j.foodchem.2020.127765
Voglino, G.F., 1972. A new β-casein variant in piedmont cattle. Anim. Genet. 3(1): 61–62. 10.1111/j.1365-2052.1972.tb01233.x