
Download
PAPER
The role of whey protein in myogenic differentiation
Rui Xu1*, Yuejuan Xiao2, Ying Zhang2, Xiyan Zhao1
1College of Food Science and Technology, Hebei Normal University of Science and Technology, Changli, Qinhuangdao, China;
2College of Marine Resources and Environment, Hebei Normal University of Science and Technology, Changli, Qinhuangdao, China
Abstract
Whey protein has been shown to prevent exercise-induced protein degradation and muscle damage. We hypothesized that whey protein would regulate muscle cell differentiation. Adding various concentrations of whey protein to C2C12 myoblasts induced cell differentiation and MyoD (myogenic differentiation protein) expression as well as the phosphorylation of AKT (protein kinase B). Whey protein-induced differentiation-specific markers increased the enzymatic activities of creatine kinase and citrate synthase and the expression of muscle-specific microRNA. Whey protein elevated AKT phosphorylation on Thr308 and Ser473, which was inhibited by LY294002 (a non- selective phosphoinositide 3-kinases inhibitor), suggesting that whey protein acts via PI3K (phosphatidylinositol 3-kinase). Blocking of the PI3K/AKT pathway with specific inhibitors revealed its requirement in mediating the promotive effects of whey protein on C2C12 cell differentiation. These effects of whey protein on myoblast differentiation suggest its positive influence in preventing muscle atrophy.
Key words: C2C12, myoblast differentiation, whey protein
*Corresponding Author: Rui Xu, College of Food Science and Technology, Hebei Normal University of Science and Technology, Changli, Qinhuangdao, 066600, China. Email: [email protected]
Received: 25 July 2021; Accepted: 28 March 2022; Published: 26 April 2022
© 2022 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Whey protein, including essential branched-chain amino acid, is one of the latest dietary supplements for promoting/aiding the increase in strength and improving the lean body mass. The branched-chain amino acid has been shown to increase muscle hypertrophy by inhibiting muscle degradation (Aoyama et al., 2019). A change in lean muscle mass is often attributed to a changed balance between protein synthesis and degradation rates in tissues. However, the direct effect of whey protein on myogenic mechanisms has never been studied. Here we examined the effects of whey protein on the differentiation of C2C12 myoblasts.
Skeletal muscle formation or myogenesis is a complex and highly regulated process that involves the proliferation of myoblasts, exit of these myoblasts from the cell cycle, and their differentiation into muscle fibers (Hernández-Hernández et al., 2017). The sequential expression of myogenic regulatory factors (MRFs), a group of basic helix-loop-helix (bHLH) transcription factors that include MyoD, Myf-5 (myogenic factor 5), myogenin, and MRF4 is required (Seale and Rudnicki, 2000). MyoD and Myf-5 are the primary MRFs required for the formation, proliferation, and survival of myoblasts whereas myogenin and MRF-4 act later to control myoblast differentiation and induce the expression of important muscle-specific genes, such as myosin heavy chain and creatine kinase.
MicroRNAs (miRNAs) are a class of highly conserved, non-coding RNAs molecules that play key roles in posttranscriptional gene regulation. A small number of muscle-specific miRNAs have been shown to have a crucial role in myoblast proliferation and differentiation (Wang et al., 2018). Numerous miRNA expression profiling studies have consistently shown that miRNA-1 (miR-1), miR-133a, and miR-206 have to be muscle-specific (Motaei et al., 2019). Analysis of the presumptive miR-1 promoter has shown that miR-1 expression is regulated by SRF (serum response factor), MyoD, MEF2 (myocyte enhancer factor-2), and Dorsal- and Twist-binding sites, all factors known to be important in conferring muscle- specific gene expression (Zhang et al., 2021). miR-1 promotes the differentiation and exit of cardiac and skeletal progenitors from the cell cycle in mammals.
The phosphoinositide 3'-kinase (PI3K) plays a crucial role in effecting alterations in a broad range of cellular functions in response to extracellular signals. Akt phosphorylates a variety of substrates involved in the regulation of key cellular functions including cell growth and survival and protein translation. The PI3K/AKT pathway has been reported to play a major role in inducing cell differentiation and hypertrophy (Rommel et al., 2001).
Given the importance of the muscle-specific miRNAs and PI3K/AKT signaling in muscle development, we must have the interest to determine what role they may have in skeletal muscle plasticity. To gain insight into the roles of whey protein in muscular hypertrophy, we analyzed the profiles of miR-1, miR-133a, and miR-206 expression as well as PI3K/AKT pathway in C2C12 cells.
Materials and Methods
Materials
C2C12 myoblast was purchased from China Center for Type Culture Collection. Dulbecco’s modified Eagle’s medium (DMEM) and fetal bovine serum (FBS) were purchased from Gibco (Grand Island, USA). Whey protein concentration contained 80.05% protein, which was measured using the Kjeldahl method. LY294002 is LY 294002 HYDROCHLORIDE, purchased from Beyotime Institute of Biotechnology (Jiangsu, China).
Cell culture
The cells were grown in DMEM containing 10% FBS. Once cells had reached 80% confluency, the medium was changed to DMEM supplemented with 2% horse serum. C2C12 myoblasts were differentiated into myotubes by incubation in the differentiation medium (2% horse serum in DMEM). Myotubes were maintained in the differentiation medium, and the medium was changed every 48 h. The inhibitors LY294002 (20 μM) was added before whey protein, which was used at 0.1 mg/mL or 0.4 mg/mL for C2C12 myoblasts.
Enzyme assays
The activities of two marker enzymes of differentiation, creatine kinase (CK) and citrate synthetase (CS), were assayed in cellular extracts using a spectrophotometric- based kit from Nanjing Jiancheng Bioengineering Institute (Nanjing, China) and Shanghai Genmed Pharmaceutical Co., Ltd (Shanghai, China), respectively.
Detection of mature miRNAs
To detect miRNA in all RNA samples, the All-in-one miRNA Q-PCR detection kit and PCR primer sets for miR-1, -133a, and -206 were used according to the manufacturer’s directions (GeneCopoeia, Guangzhou, China). Further, the expression of all samples was normalized to Rnu6 (U6 small nuclear RNA) levels to account for possible differences in the amount of initial RNA.
Quantitative real-time PCR
Isolation of all RNA and RT-PCR was done as described by Dogra et al. (2006), but with a minor modification. Briefly, an RNAiso Plus Kit (TaKaRa, Dalian, China) was used to extract RNA from cells. All RNA samples were reverse-transcribed using gene-specific primer and Fermentas’s reverse transcription kit according to the manufacturers’ instructions. The sequences of primers used were as follows: MyoD, 5'-TGCGGTGCACCCAGGCCCAG-3' (forward) and 5'-CCGCCTCACTGTAGTAGGCG-3' (reverse); Myf5, 5'-CAAGAGTAGCAGCCTTCGGA-3' (forward) and 5'-GGAGCTTTTATCTGCAGCAC-3'(reverse); Myogenin, 5'-GCTATGAGCGGACTGAGCTC-3' (forward) and 5'-GGAGTGCAGATTGTGGGCGT-3' (reverse); GAPDH, 5'-CCTTCATTGACCTCAACTAC-3' (forward) and 5'-AGCCCCACGGCCATCACGCC-3' (reverse).
Quantitative real-time PCR was conducted using the same primers as described above and following a method as detailed by Dogra et al. (2006). Data normalization was accomplished using the endogenous control (GAPDH) and the normalized values were subjected to a 2–ΔΔCt threshold cycle formula to calculate the fold change between the control and experiment groups.
Western blot analysis
Cells were extracted with (Radio Immunoprecipitation Assay) (RIPA) buffer and lysates were cleared by centrifugation. Equal amounts of protein were loaded and resolved by 12% (w/v) SDS-PAGE, then transferred to nitrocellulose membranes (Whatman). The membranes were blocked with 5% nonfat dry milk in Trisbuffered saline-0.1% Tween (TBST) and probed overnight with primary antibodies in TBST supplemented with 5% bovine serum albumin. After reaction in TBST with horseradish peroxidase-conjugated secondary antibodies (Boisynthesis, Beijing, China), bands were visualized using enhanced HRP-DAB reagents (Tiangen, Beijing). After blocking, the membranes were incubated with the following primary antibodies: polyclonal anti-AKT (Cell Signaling), anti-phospho-AKT (Thr308), anti-phospho-AKT (Ser473), and monoclonal anti-myosin heavy chain (Santa Cruz), and polyclonal anti-β-Actin antibody (Boisynthesis).
Statistical analysis
Results are expressed as mean ± SE. A value of P < 0.05 was considered statistically significant unless otherwise specified and statistical analyses were performed using Origin 8.0.
Results
Assay of marker enzyme
The addition of various concentrations of whey protein to serum-starved C2C12 myoblasts for 24 h enhanced the enzymatic activity of the muscle differentiation-specific markers creatine kinase and citrate synthase in a dose-responsive manner, with the highest levels being observed at 0.4 mg/ml whey protein. The enzyme activities of CS and CK are shown in Table 1. There was no significant change in CS activities for the concentration of 0.1 mg/ml. Comparisons of the activities of markers of differentiated skeletal muscle CS and CK revealed increases from 2.61 to 5.87 U/mg·prot and from 41.7 to 68.4 μmol/mg·min, respectively.
Table 1. Enzymatic activity of two marker enzymes of differentiation.
| Activity | Control | 0.1 mg/ml whey | 0.4 mg/ml whey |
|---|---|---|---|
| CK(U/mg·prot) | 2.61 ± 0.12 | 3.94 ± 0.25* | 5.87 ± 0.39+ |
| CS(μmol/mg·min) | 41.7 ± 3.09 | 53.2 ± 3.22 | 68.4 ± 4.41* |
Data are expressed as means ± SE.
*Statistically significant difference from Control (P < 0.05).
+ Significantly different from Control (P < 0.01).
Myogenic gene expression
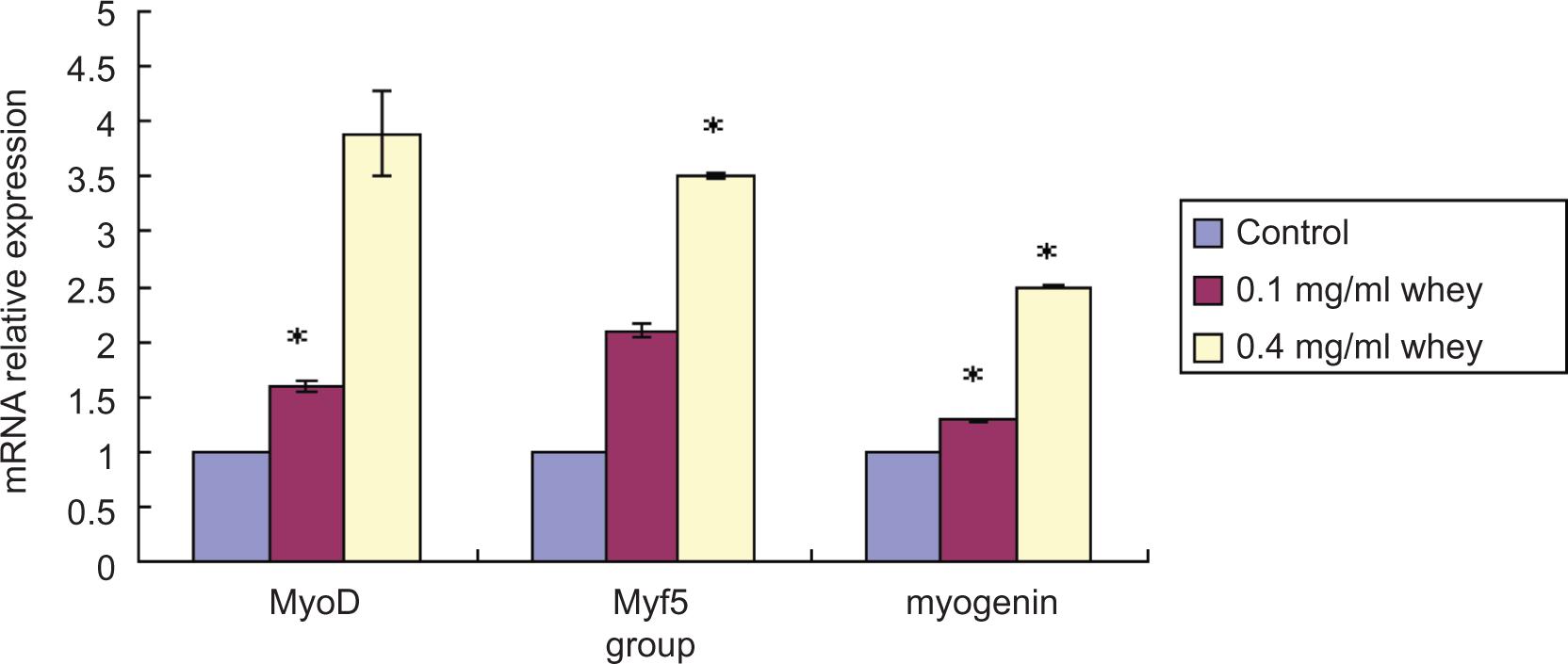
Since the activation of MRFs is a prerequisite for myogenesis, we investigated the effect of whey protein on the expression of MyoD, myogenin, and Myf-5. C2C12 myoblasts were incubated in DM alone or with 0.1 and 0.4 mg/ml whey protein. The expression of MyoD, myogenin, and Myf-5 mRNA was measured using QRT-PCR. As shown in Figure 1, whey protein increased the expression of MyoD, Myf5, and Myogenin in C2C12 myoblasts. The mRNA levels of these genes were significantly higher in the experimental group as compared with the control group. These data suggest that whey protein regulates myogenic differentiation by influencing the MRF transcript levels.
Figure 1. Whey protein increases markers of myogenic differentiation. C2C12 myoblasts were incubated in the absence or presence of whey protein at various concentrations for 24 h, and mRNA expression levels of myogenin, MyoD, and Myf5 were analyzed by quantitative real-time PCR using GAPDH as an internal control. *Statistically significant difference from Control (P < 0.05).
miRNA expression
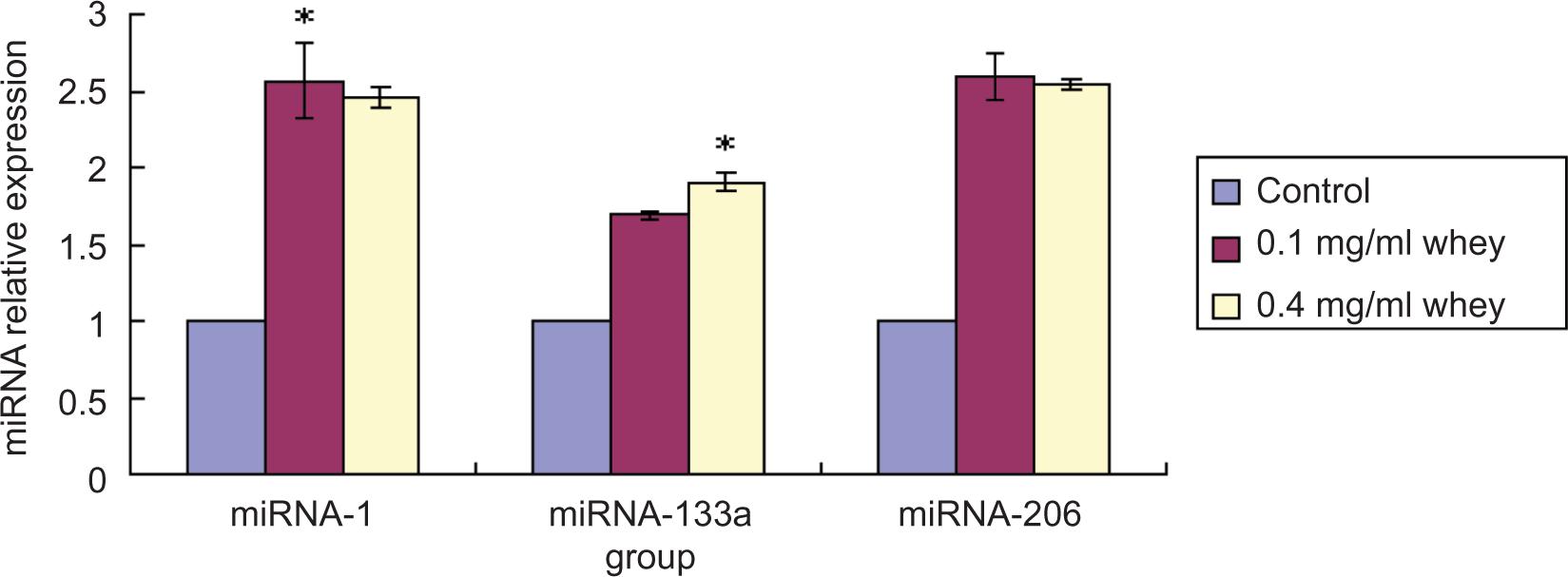
There was no difference in the level of expression for miRNA-206, which was 2.6-fold higher in the low-dose group relative to the control group. As shown in Figure 2, expression of miRNA-1 and miRNA-206 increased by ~2.5-fold in response to whey protein, whereas miRNA-133a expression increased by ~1.7-fold. In contrast to the other miRNA, miRNA-206 and miR-133a expression did not change significantly after the low-dose treatment. The observed change in miRNA expression suggested there would be a similar change in the expression of the mature muscle-specific miRNAs, miR-1, miR-133a, and miR-206.
Figure 2. Whey protein increases the expression of muscle-specific miRNA. C2C12 myoblasts were incubated in the absence or presence of whey protein at various concentrations for 24 h, and miRNA expression levels were analyzed by fluorescence quantitative real-time PCR using U6 as an internal control. *Statistically significant difference from Control (P < 0.05).
PI3K/AKT signaling pathways
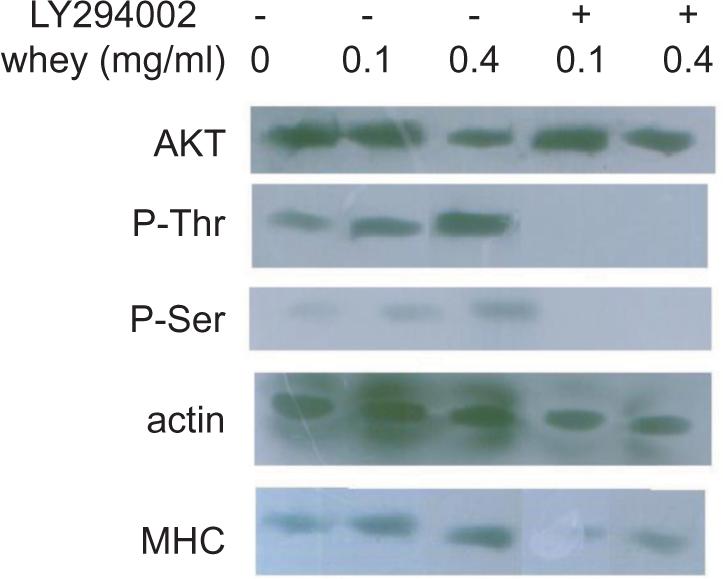
Given our results, we sought to determine whether whey protein induces signaling pathways such as that of PI3K/AKT. In serum-starved C2C12 cells, AKT phosphorylation on amino acid Ser473 was upregulated after incubation with whey protein in an elevated manner. AKT phosphorylation was induced in C2C12 cells in a dose-dependent manner (Figure 3). Whey protein induced the phosphorylation of AKT phosphorylation sites, Ser473 and Thr308, both are required for full kinase activity. This phosphorylation was completely abolished in the presence of the specific PI3K inhibitor LY294002. LY294002 treatment completely abolished the whey-induced AKT phosphorylation, implying the requirement of PI3K activation for this response. Increases in protein synthesis require activation of the translational machinery and in addition may consist of indirect effects after stimulation of gene transcription. To test whether AKT phosphorylation is required for whey protein’s stimulatory effect on myoblast protein synthesis, C2C12 myoblasts were treated with 0.1 and 0.4 mg/ml whey protein for 24 h with or without Ly294002, a stable PI3K inhibitor, which was added 30 min before the whey protein addition. A western blot analysis for MHC (myosin heavy chain) revealed that Ly294002 addition prevented the whey-induced MHC levels in C2C12 cells (Figure 3).
In the absence of LY294002, treatment with 0.4 mg/ml whey protein increased baseline protein synthesis compared to untreated controls for Serine and Threonine phosphorylation, respectively. These results indicate a strong impact of PI3K on baseline protein synthesis. The whey-induced increase in protein synthesis was reduced by LY294002 above the corresponding control conditions, indicating that PI3K contributes to whey protein-induced protein synthesis.
The whey-induced PI3K-dependent AKT phosphorylation is required for whey’s promotive effect on the differentiation of myoblasts. Taken together, these data suggest that whey protein has a beneficial effect on adult skeletal muscle, both during normal growth and under stress conditions.
Discussion
Our study demonstrates that whey protein promotes general protein accretion, muscle-specific gene expression, and myoblast fusion during myogenic differentiation. Moreover, our data suggest that stimulation of these processes by whey protein relies on positive regulation of AKT because increased AKT phosphorylation and a subsequent increase in MHC protein were observed in response to whey protein. In the enzymatic assay of CK and CS, the well-known and accepted markers of differentiation, cultures treated with a differentiation medium showed a higher activity, evidencing a higher degree of differentiation. As expected, a dose-dependent promotion in both CK and CS activity was seen in the presence of whey protein in C2C12 cultures. Many literatures (Griffen et al., 2022; Thalacker-Mercer et al., 2020) have shown that whey protein can promote muscle hypertrophy because it contains branched-chain amino acids. In the pre-experiment, we found that when whey protein concentration was 0.1 mg/ml, it showed an effect compared with the control group, so we chose these two doses (0.1 and 0.4 mg/ml).
Skeletal muscle differentiation can be regulated by at least three possible mechanisms, which include alteration in protein degradation, mRNA stabilization, and gene transcription. Although evidence exists to support the first two mechanisms (Lee et al., 2019; Vaisid and Kosower, 2013), modulation in gene expression appears to be most important during differentiation. This occurs via the activation of various phosphatases and kinases, which in turn alter the activity of downstream regulatory factors such as MEF2, Myf-5, myogenin, MyoD, serum response factor, and nuclear factor of activated T cells. Since the presence of whey protein increased the transcripts of MyoD, myf-5, and myogenin (Figure 1) and also promoted the protein synthesis of MHC in myoblasts (Figure 3), our data suggest that the increased myogenesis is the result of increased transcription of MRFs.
Figure 3. The PI3K/Akt pathway is induced by whey protein. Akt phosphorylation of serum-starved C2C12 myoblasts in response to various concentrations of whey protein. Akt phosphorylation was analyzed by western blot with the anti-phospho-Akt antibody followed by densitometric analysis normalized to levels of β-actin. The western blot analysis for MHC in C2C12 cells treated for 24 h with different concentrations of whey protein without or with LY294002 (25 μM).
The current investigation was undertaken as an initial survey of muscle-specific miRNA expression in response to whey-induced differentiation to determine if these novel trans-factors have a potential role in adult skeletal muscle hypertrophy. The results of this study comprehensively provide the first evidence of miRNAs functions during whey-induced muscle hypertrophy and the foundation for future studies directed at elucidating the role of specific miRNAs during muscle growth. The possibility that miR-206 may have a role in the regulation of skeletal muscle hypertrophy is supported by a recent study demonstrating that miR-206 contributes to the hypertrophic phenotype of Texel sheep (Qin et al., 2017). Quantitative trait loci mapping revealed Texel sheep have a single nucleotide polymorphism (SNP) within the 3'-UTR of the myostatin gene that results in the formation of a functional miR-1/miR-206 target site.
miR-1, miR-133a, and miR-206 are transcribed during myogenesis and seem to be regulated by MyoD and myogenin (Rosenberg et al., 2006). miR-206 is transcribed independently of miR-1 and miR-133a, which are transcribed as a common pri-miRNA precursor from the miR-1/miR-133a locus, followed by alternative splicing to generate different primary transcripts (Taylor and Hughes, 2017). This is consistent with the present finding that miR-1 and miR-133a expression showed similar patterns in their induction and increase during differentiation. These observations suggested that miR-206 might be induced by MyoD whereas miR-1 and miR-133a might be induced by myogenin, as the onset and temporal sequence of miR-206 and miR-1/miR-133a expression seem to coincide with those of MyoD and myogenin expression during myogenesis. After their induction by MyoD and myogenin, however, other factors must be involved in the expression of these miRNAs at later stages.
The challenge for future studies will be to identify relevant target genes of each muscle-specific miRNA and how they contribute to the regulation of skeletal muscle growth and phenotype. Further studies are needed to address these complex regulatory roles of miRNAs, and the identification of bona fide, biologically relevant targets for miRNAs will be an important goal for miRNA researchers in the future.
Whey protein has been observed to influence muscle strength via various mechanisms, such as decreasing protein degradation and attenuating the proteosome–ubiquitin degradation pathway on the one hand, and increasing protein synthesis on the other. Taken together, our findings suggest that, at least in culture, whey protein can drive C2C12 myoblasts into the cell cycle, thus acting as a mitogen in these cells. The differential response of C2C12 cells to whey protein suggests this mitogen’s species-specific effect. Many investigators have reported the stimulatory effect of essential amino acids on mRNA translation (Sans et al., 2021), which generally relies on the enhanced activity of the ribosomal protein S6 kinase (S6K1) resulting from signaling through the PI3K/Akt/mTOR pathway.
Whey protein increased the protein levels of MHC in a dose-dependent manner. The signaling pathways through which whey protein exerts its effects are not fully understood. In this study, we demonstrate for the first time the promotive effect of whey protein on AKT phosphorylation and the requirement of this pathway to mediate its effects, as demonstrated by employing specific inhibitors. By blocking the PI3K/AKT pathway with specific inhibitors, we demonstrate that this pathway is required in mediating the promotive effect of whey protein on muscle cell differentiation. The crucial role of AKT in myogenic differentiation and hypertrophy induced by whey protein has been well demonstrated in this study.
Recent studies have reported that a class III PI3K mediates the amino acid activation of the mammalian target of rapamycin (mTOR) and its downstream molecules, which is distinct from the effect of insulin (Kwak et al., 2016). Our findings indicate that whey protein fully activates AKT by inducing phosphorylation on both Thr308 and Ser473, while the latter phosphorylation is inhibited by LY294002, a specific PI3K inhibitor. However, we cannot rule out the possibility that whey protein activates the mTOR/p70S6k pathway in a manner similar to other amino acids.
Therefore, we believe that the promotive effects, shown here, of whey protein on muscle cell differentiation, together with its previously demonstrated anti-catabolic effects, justify its supplement as a therapeutic strategy to prevent muscle loss in myopathies as well as in aging and trauma. However, together with mRNA content, it could be expressed by showing the differences in cell morphology and quantifying the level of index fusion of the number of nuclei by another measurement method. Moreover, the PI3K/AKT axis such as p38 MAPK could also be discussed and we would have a great interest in it in the future.
Acknowledgments
This project was sponsored by the research project of Hebei Education Department, China (ZC2021230).
Conflict of Interest
There are no conflicts of interest.
REFERENCES
Aoyama, S., Hirooka, R., Shimoda, T. and Shibata, S., 2019. Effect of different sources of dietary protein on muscle hypertrophy in functionally overloaded mice. Biochemistry and Biophysics Reports. 20: 100686–100693.
Dogra, C., Changotra, H., Mohan, S. and Kumar, A., 2006. Tumor necrosis factor-like weak inducer of apoptosis inhibits skeletal myogenesis through sustained activation of nuclear factor-kappaB and degradation of MyoD protein. Journal of Biological Chemistry 281: 10327–10336. 10.1074/jbc.M511131200
Griffen, C., Duncan, M., Hattersley, J., Weickert, M.O., Dallaway, A. and Renshaw, D., 2022. Effects of resistance exercise and whey protein supplementation on skeletal muscle strength, mass, physical function, and hormonal and inflammatory biomarkers in healthy active older men: a randomised, double-blind, placebo-controlled trial. Experimental Gerontology 158: 111651–111657. 10.1016/j.exger.2021.111651
Hernández-Hernández, J.M., García-González, E.G., Brun, C.E. and Rudnicki, M.A., 2017. The myogenic regulatory factors, determinants of muscle development, cell identity and regeneration. Seminars in Cell & Developmental Biology 72: 10–18. 10.1016/j.semcdb.2017.11.010
Kwak, S.S., Kang, K.H., Kim, S., Lee, S., Lee, J.-H., Kim, J.W., Byun, B., Meadows, G.G. and Joe, C.O. et al, 2016. Amino acid-dependent NPRL2 interaction with Raptor determines mTOR Complex 1 activation. Cellular Signalling 28(2): 32–41. 10.1016/j.cellsig.2015.11.008
Lee, S.-J., Bae, J.H., Lee, H., Lee, H., Park, J., Kang, J.-S. and Bae, G.-U., 2019. Ginsenoside Rg3 upregulates myotube formation and mitochondrial function, thereby protecting myotube atrophy induced by tumor necrosis factor-alpha. Journal of Ethnopharmacology 242: 112054–112059. 10.1016/j.jep.2019.112054
Motaei, J., Yaghmaie, M., Ahmadvand, M., Pashaiefar, H. and Kerachian, M.A., 2019. MicroRNAs as potential diagnostic, prognostic, and predictive biomarkers for acute graft-versus-host disease. Biology of Blood and Marrow Transplantation 25(12): e375–e386. 10.1016/j.bbmt.2019.08.004
Qin, Y., Peng, Y., Zhao, W., Pan, J., Ksiezak-Reding, H., Cardozo, C., Wu, Y., Pajevic, P.D., Bonewald, L.F., Bauman, W.A. and Qin, W. et al, 2017. Myostatin inhibits osteoblastic differentiation by suppressing osteocyte-derived exosomal microRNA-218: a novel mechanism in muscle-bone communication. Journal of Biological Chemistry 292(26): 11021–11033. 10.1074/jbc.M116.770941
Rommel, C., Bodine, S.C., Clarke, B.A., Rossman, R., Nunez, L., Stitt, T.N., Yancopoulos, G.D. and Glass, D.J., 2001. Mediation of IGF-1-induced skeletal myotube hypertrophy by PI(3)K/Akt/mTOR and PI(3)K/Akt/GSK3 pathways. Nature Cell Biology 3: 1009–1013. 10.1038/ncb1101-1009
Rosenberg, M.I., Georges, S.A., Asawachaicharn, A., Analau, E. and Tapscott, S.J., 2006. MyoD inhibits Fstl1 and Utrn expression by inducing transcription of miR-206. Journal of Cell Biology 175: 77–85. 10.1083/jcb.200603039
Sans, M.D., Crozier, S.J., Vogel, N.L., D’Alecy, L.G. and Williams, J.A., 2021. Dietary protein and amino acid deficiency inhibit pancreatic digestive enzyme mRNA translation by multiple mechanisms. Cellular and Molecular Gastroenterology and Hepatology 11(1): 99–115. 10.1016/j.jcmgh.2020.07.008
Seale, P. and Rudnicki, M.A., 2000. A new look at the origin, function, and “stem-cell” status of muscle satellite cells. Developmental Biology 218: 115–124. 10.1006/dbio.1999.9565
Taylor, M.V. and Hughes, S.M., 2017. Mef2 and the skeletal muscle differentiation program. Seminars in Cell & Developmental Biology 72: 33–44. 10.1016/j.semcdb.2017.11.020
Thalacker-Mercer, A., Riddle, E. and Barre, L., 2020. Chapter two–protein and amino acids for skeletal muscle health in aging. Advances in Food and Nutrition Research 91: 29–64. 10.1016/bs.afnr.2019.08.002
Vaisid, T. and Kosower, N.S., 2013. Calpastatin is upregulated in non-immune neuronal cells via toll-like receptor 2 (TLR2) pathways by lipid-containing agonists. Biochimica et Biophysica Acta (BBA)–Molecular Cell Research 1833(10): 2369–2377. 10.1016/j.bbamcr.2013.06.006
Wang, J., Yang, L.Z., Zhang, J.S., Gong, J.X., Wang, Y.H., Zhang, C.L., Chen, H. and Fang, X.T., 2018. Effects of microRNAs on skeletal muscle development. Gene 668: 107–113. 10.1016/j.gene.2018.05.039
Zhang, H., Wang, Y., Tang, X., Dou, S., Sun, Y., Zhang, Q. and Lu, J., 2021. Combinatorial regulation of gene expression by uORFs and microRNAs in Drosophila. Science Bulletin 66(3): 225–228. 10.1016/j.scib.2020.10.012