
Download
ORIGINAL ARTICLE
Unveiling the anti-tumor and anti-fibrotic potentials of Kanglaite in esophageal cancer through PVT1/TGF-β/Smad axis inhibition
Xiuqing Li1, Ka Bian2, Ji Zhang3, Kaili Luo1, Tao Dong1, Chenglin Luo3*
1Department of Gastroenterology and Hepatology, The Third People’s Hospital of Zhenjiang, Zhenjiang, Jiangsu, China;
2Department of Gastroenterology and Hepatology, Yangzhou University Medical College, Yangzhou, Jiangsu, China;
3Department of Gastroenterology and Hepatology, Suzhou Xiangcheng People’s Hospital, Suzhou, Jiangsu, China
Abstract
Esophageal cancer (EC) poses a significant challenge among tract tumors worldwide due to its complex molecular composition, which remains largely unexplored. Kanglaite (KLT), a medicine known for its anti-tumor properties, is emerging as a potential ally in the culinary world. Plasmacytoma Variant translocation 1 (PVT1) plays a significant role in the complex nature of esophageal cancer, showing high expression levels in tumor tissues. During our investigation, we discovered a higher presence of PVT1 in cancer tissues, which led us to explore its functions further. Through manipulating techniques, we found that inhibiting PVT1 not only hinders the growth of esophageal cancer cells but also reduces fibrosis. This is similar to enhancing the texture and flavor of a dish. Our study reveals the interactions at play, highlighting how PVT1 orchestrates the TGF β/Smad pathway – a component in the development of esophageal cancer. Introducing KLT, an expert poised to revolutinize the approach to cancer treatment. Our research demonstrates KLT’s effectiveness in regulating the PVT1/TGF-β/Smad axis, uncovering its fibrotic properties, and its ability to hinder the progression of esophageal cancer. With these discoveries, KLT emerges as a promising option for the treatment of esophageal cancer.
Key words: esophageal cancer, fibrosis, Kanglaite, plasmacytoma variant translocation 1, TGF-β/Smad pathway
*Corresponding Author: Chenglin Luo, Department of Gastroenterology and Hepatology, Suzhou Xiangcheng People’s Hospital, No. 1060 Huayuan Road, Xiangcheng District, Suzhou, Jiangsu, 215100, China. Email: [email protected]
Received: 8 May 2024; Accepted: 10 June 2024; Published: 1 July 2024
© 2024 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Esophageal cancer (EC) is a global issue that significantly impacts the culinary world, particularly prevalent in China, where it ranks among the top five cancers. In the array of food choices, esophageal cancer poses a challenge that demands increased attention and creative culinary solutions to reduce its harmful effects (Yang et al., 2020). Every year, this type of cancer takes a toll on cuisine, claiming about 300,000 lives. This widespread disease raises concernsand highlights the need for serious culinary strategies to mitigate its impact on human health (Sun et al., 2023).
Its morbidity and mortality rates vary widely (Yanagida et al., 2024). In the realm of health issues, China stands out as a significant area with a high rate of esophageal cancer cases. This country, known for its cuisine, is facing a significant challenge with this type of cancer. As a result, it becomes important to focus on culinary studies and efforts to reduce its negative effects on public health (Lian et al., 2023). Esophageal squamous cell carcinoma (ESCC) is the main EC type (Kashiwagi et al., 2023). High invasiveness is one of the characteristics of EC (Kashiwagi et al., 2023; Liu et al., 2022; Wang et al., 2021). The survival rate of EC patients is poor due to local infiltration (Kashiwagi et al., 2023). However, the molecular mechanisms that cause EC’s malignant behavior are not fully understood.
Plasmacytoma Variant translocation 1 (PVT1) is an organizer positioned downstream of human chromosome 8q24, captivating tumor tissues with its expression levels (Dong et al., 2023; Luo et al., 2022; Sun et al., 2023). High PVT1 expression in these tumors indicates a poor prognosis (Dong et al., 2023; Luo et al., 2022; Sun et al., 2023). Similar to how a skilled chef combines ingredients to create a dish, PVT1 manipulates the components in pancreatic cancer cells, developing resistance to gemcitabine, a crucial element in cancer treatment. PVT1’s impact could be compared to regulating the Wnt/β pathway, similar to controlling heat levels for the simmer. Exploring the complexities of cancer resistance reveals PVT1’s expertise as a master chemist, highlighting potential targets for therapeutic strategies in combating cancer (Wu et al., 2017). Our findings are consistent with previous studies demonstrating the role of PVT1 in cancer progression and fibrosis. For example, various studies have shown that PVT1 enhances tumor growth and promotes resistance to chemotherapy in different types of cancers, such as gastric and pancreatic cancers. Moreover, Kanglaite’s ability to inhibit the PVT1/TGF-β/Smad pathway supports its efficacy in reversing drug resistance and suppressing tumor growth through various signaling pathways, in line with other findings.
The PVT1 gene plays a key role in esophageal cancer (EC). Studies have indicated a connection between PVT1 levels and specific traits, highlighting its significant influence on EC development. PVT1 controls the growth and movement of EC cells by adjusting the miR 128/ZEB1 axis. This intricate orchestration is directed by PVT1, guiding the actions that drive the progression of EC (Zheng et al., 2016). PVT1 also promotes TGF-β/Smad pathway activation, thus contributing to tumor progression (Li et al., 2022).
Kanglaite (KLT) has been reported to have antitumor activity (Lv et al., 2023; Zhao et al., 2023). KLT is an oily substance of coix seed (Zhao et al., 2023). Its main active ingredient is a triglyceride compound containing four fatty acids. Kanglaite suppressed the drug resistance gene expression by inhibiting PVT1 in cisplatin-resistant gastric cancer cells (Zhang et al., 2017). Therefore, we speculated that the antitumor effect of Kanglaite was related to the inhibition of PVT1.
Despite the promising results, our study has several limitations. First, the research was primarily conducted in vitro, and the in vivo efficacy and safety of KLT need to be further validated. Second, further investigation is needed to understand the KLT’s inhibition of PVT1 and the TGF-β/Smad pathway at the molecular level. Third, our sample size was relatively small, and larger studies are needed to confirm these findings. Future studies should prioritize conducting comprehensive in vivo experiments to evaluate the therapeutic potential of KLT in animal models of esophageal cancer. Additionally, exploring the detailed molecular interactions between KLT, PVT1, and the TGF-β/Smad pathway will provide deeper insights into its mechanism of action. Clinical trials are also recommended to determine the efficacy and safety of KLT in patients with esophageal cancer.
Our research aimed to explore the cooking potential of KLT for cancer. We found an increase in the expression of PVT1 in cancer tissues and uncovered its effects on EC progression. KLT exerts its anticancer effects by downregulating the expression of PVT1, thereby inhibiting the TGF-β/Smad pathway, which is crucial for tumor progression and fibrosis in esophageal cancer. This regulatory mechanism underscores KLT’s potential in controlling esophageal cancer growth and fibrotic responses.
Materials and Methods
Samples
The 30 EC tissues and 30 corresponding normal tissues were collected from the patient’s surgical therapy in The Third People’s Hospital of Zhenjiang. The ethics committee of the Third People’s Hospital of Zhenjiang approved the procedures for the current study.
Cell culture and transfection
Human esophageal epithelial cell line SHEE and four types of EC cells, including KYSE-30, KYSE-450, KYSE-150, and KYSE-510 cells, were all purchased from the Cell Bank of the Chinese Academy of Sciences. Cells were cultured with the complete medium (DMEM medium containing 10% FBS). After 24 h of culture, Lipofectamine®3000 reagent (Invitrogen, USA) was co-transfected with sh-NC, sh-PVT1, pcDNA3.1-vector, and pcDNA3.1-PVT1, respectively. The sequences of shRNA were shown as follows: sh-NC: 5’-UUCUCCGAACGUGUCACGUTT-3’; sh-PVT1: 5’-GGAAGTCTTGCTTCTTTAT-3’. 100 μL of the mixture was slowly added to the 6-well plate. After 24 h of culture, cells were treated with KLT (bought from Sigma) for 24 h at concentrations of 0, 1, 2.5, and 5 μL/mL. Then, the effect of KLT was verified for subsequent experiments.
qPCR
TRIzol® (cat. no. 15596026; Invitrogen; Thermo Fisher Scientific, Inc.) reagent was used to isolate RNA from HUVECs. Total RNA was reverse transcribed into cDNA at 42°C for 1 h using M-MLV reverse transcriptase (cat. no. M1701; Promega Corporation), which consisted of the following: 5 µL M-MLV 5X Reaction Buffer, 1.25 µL dNTP (10 mM), 25 units of Recombinant RNasin® Ribonuclease Inhibitor, 200 units of M-MLV RT, and nuclease-free water to a final volume of 25 µL. qPCR was performed using the kit TB Green Premix Ex Taq (Tli RNase H Plus) [cat. no. RR420A; Takara Bio, Inc.] under the following thermocycling conditions: initial denaturation at 95°C for 3 min; followed by 30 cycles of denaturation at 95°C for 30 sec, annealing at 58°C for 30 sec and extension at 72°C for 30 sec; and a final extension step at 4°C for 30 min. The 2-ΔΔCq method was used to quantify the results (Kashiwagi et al., 2023). The sequences were shown as follows: PVT1: 5’-ATGGGACCTGTTAGTTGTG-3’, 5’-AGTCTGTCTCTGTGGCTT-3; β-actin: 5’-ACTGGAACGGTGAAGGTGAC-3’, 5’-AGAGAAGTGGGGTGGCTTTT-3’.
Western blotting
Cells were lysed in a buffer containing 1% Triton X-100, 150 mM NaCl, and 50 mM Tris (pH 7.5). The BCA assay method was used for protein concentration determination, after which proteins were separated (20 µg/lane) by SDS-PAGE on 8% gels. Then the membranes were further transferred to the PVDF membrane. The proteins were blocked with TBST containing 5% milk for 1 h, and then the corresponding primary antibodies were added and incubated at 4°C overnight. Primary antibodies TGF-β1 (Abcam, ab215715; 1:500), α-SMA (Abcam, ab7817; 1:1000), COL1A1 (Abcam, ab138492; 1:1000), Sma2 (Abcam, ab280888; 1:1000), p-Sma2 (Abcam, ab300079; 1:1000), Sma3 (Abcam, ab40854; 1:500), p-Sma3 (Abcam, ab52903; 1:1000), β-actin (Abcam, ab8226; 1:3000), and then secondary antibodies were incubated for 1 h and photographed after chemiluminescence. The reagents used in this were purchased from Wuhan Google Co., LTD. The blots were analyzed using ImageJ 9.0 software (National Institutes of Health) (Liu et al., 2022).
CCK-8 assay
Esophageal cancer cells were plated into 96-well plates with 1000 cell density and maintained for 48 h. Cells were subsequently incubated with CCK-8 (Beyotime, Beijing, China) for 4 h. Then, the OD value was measured at 450 nm wavelength (Wang et al., 2021).
EdU assay
According to the operation instructions, EC cells were incubated with the EdU kit (Beyotime, Beijing, China). Then, cells were fixed with 5% PFA. After this, the cells were counterstained with DAPI, and images were captured using a fluorescence microscope.
Statistics
GraphPad 5.0 software was used and performed for the statistical analysis. The Student’s t-test was used to determine the statistical significance between the two groups. One-way ANOVA followed by Tukey’s post hoc test was used for multiple comparisons. Data were represented as mean ± SD, and p<0.05 was considered significant.
Results
PVT1 was highly expressed in EC tissues
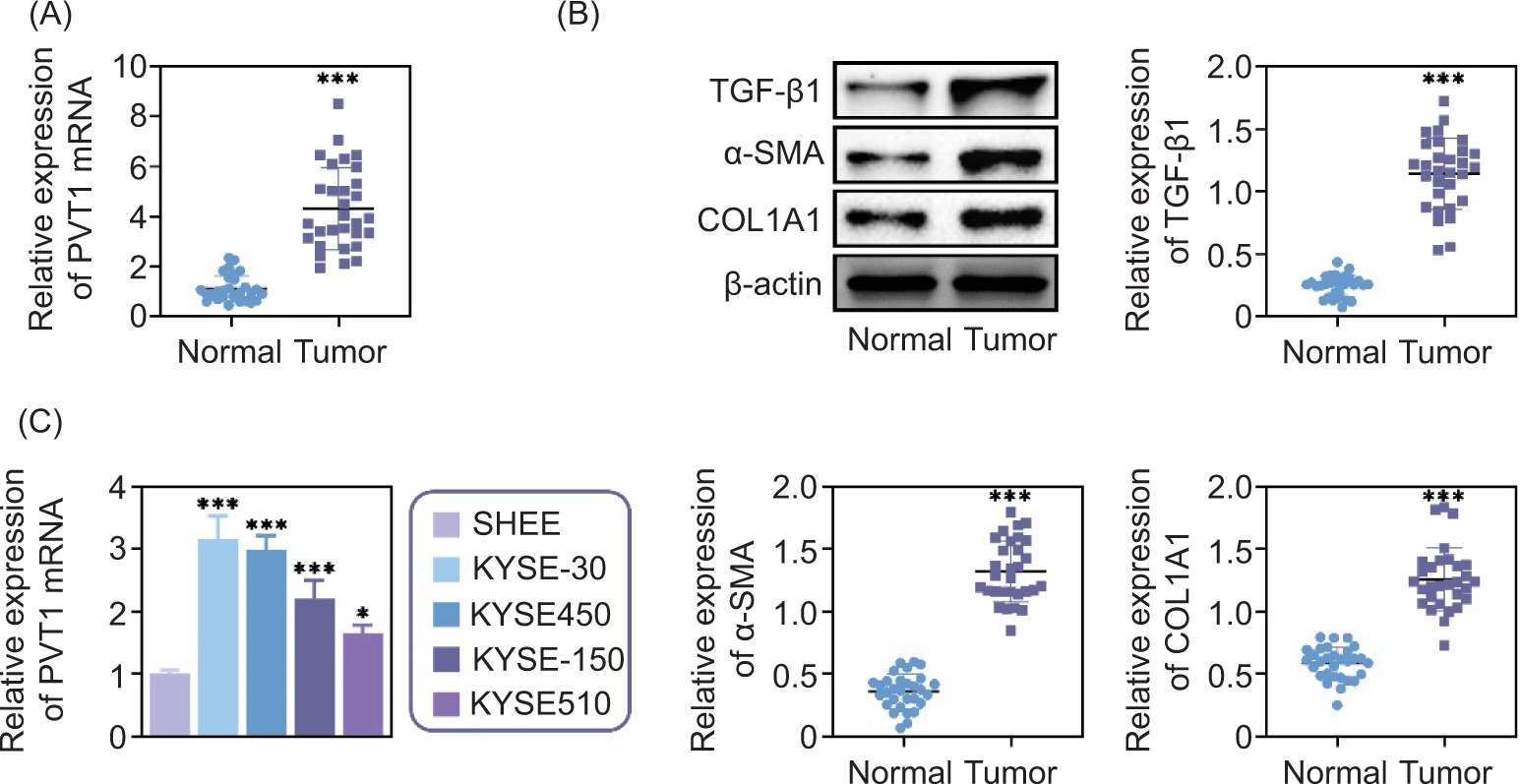
We investigated the impact of PVT1 on the advancement of cancer through qPCR analysis, comparing its mRNA levels in EC tissues with those in tissue samples. Our results revealed increased PVT1 mRNA levels in EC tissues, as illustrated in Figure 1A. Furthermore, we examined the expression levels of fibrosis markers such as TGF-β1, SMA, and COl1A1. We observed a rise in the presence of these proteins in EC tissues, indicating a fibrotic environment surroundingthe tumor (Figure 1B).
Figure 1. PVT1 was highly expressed in EC tissues. (A) qPCR assays showed the mRNA levels of PVT1 in normal as well as tumor tissues. (B) Immunoblot assays showed the expression of TGF-β1, α-SMA, and COl1A1 in normal as well as tumor tissues. The relative expression was quantified. (C) qPCR showed the mRNA levels of PVT1 in normal esophageal cell line SHEE, and EC cell line KYSE-30, KYSE-450, KYSE-150, and KYSE-510. Error bars indicate SD. *p<0.05, ***p<0.001.
We extended our study to evaluate the levels of PVT1 expression in cell types such as the SHEE esophageal epithelial cell line and four varieties of esophageal cancer cells: KYSE-30, KYSE-450, KYSE-150, and KYSE-510. Through qPCR tests, we verified the increased presence of PVT1 in all cancer cell lines, further establishing its association with cancer progression (Figure 1C). Therefore, PVT1 was found to be highly expressed in human EC.
Knockdown of PVT1 inhibited EC cell growth as well as fibrosis
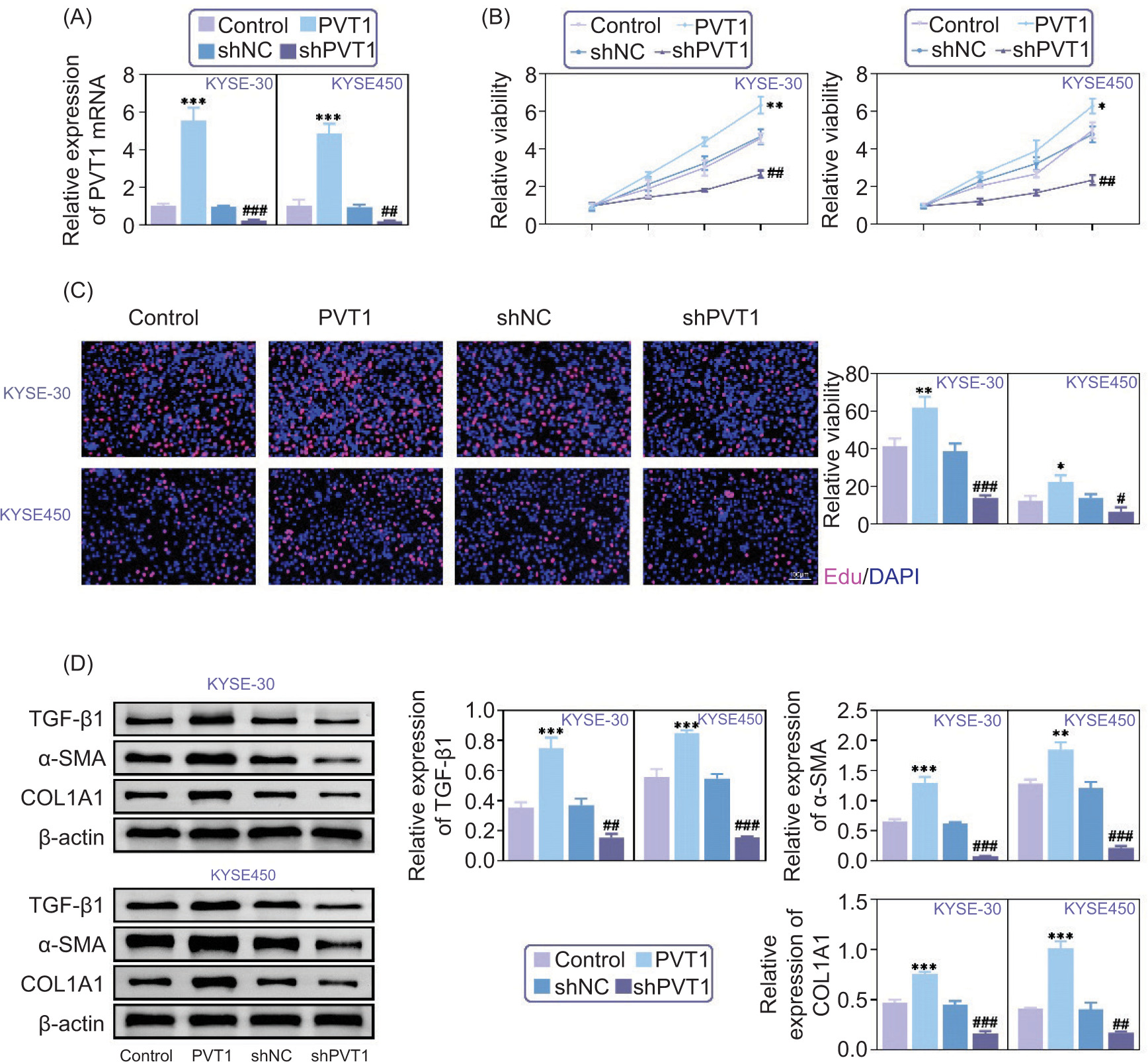
While exploring the impact of PVT1 on cancer cell behavior, we used methods to lower or raise PVT1 levels in KYSE-30 and KYSE-450 cell lines. Our detailed experiments showed that decreasing PVT1 expression with plasmids resulted in a drop in its levels while increasing its expression led to a noticeable rise, as shown in Figure 2A. Following these experiments, we examined the impact of PVT1 level variations on the growth and proliferation of cancer cells. Using CCK-8 assays, we found that decreased PVT1 levels led to reduced cell viability, as indicated by lower OD450 values (Figure 2B. Conversely, increasing PVT1 levels increased cell viability, indicating its involvement in promoting the proliferation of these cells. Additional validation through EdU assays supported these findings, showing a decrease in EdU cells when PVT1 was reduced and an increase when it was elevated (Figure 2C). This highlights the role of PVT1 in driving the proliferation of EC cells.
Figure 2. Knockdown of PVT1 inhibited EC cell growth as well as fibrosis. (A) qPCR assays showed the mRNA levels of PVT1 in KYSE-30 and KYSE-450 cells upon the indicated transfection. (B) CCK-8 assays showed the relative viability of KYSE-30 and KYSE-450 cells upon the indicated transfection. The OD450 value was quantified. (C) EdU assays showed the viability of KYSE-30 and KYSE-450 cells upon the indicated transfection. The EdU-positive cells were quantified. (D) Immunoblot assays showed the expression of TGF-β1, α-SMA, and COl1A1 in KYSE-30 and KYSE-450 cells upon the indicated transfection. The relative expression was quantified. Error bars indicate SD. *p<0.05, **p<0.01, ***p<0.001, PVT1 versus Control, #p<0.05, ##p<0.01, ###p<0.001, shPVT1 versus shNC.
To understand the interaction between PVT1 and fibrosis-related factors, we performed immunoblot assays in cancer cells to measure TGF-β1, SMA, and COl1A1 levels. Surprisingly, we found that when PVT1 levels decreased, the expression of fibrosis factor also decreased. On the other hand, when PVT1 was overexpressed, these factors were increased (Figure 2D), indicating that PVT1 plays a significant role in promoting fibrosis in KYSE-30 and KYSE-450 cells. In summary, our findings suggest that reducing PVT1 not only slows down EC cell growth but also reduces fibrosis, highlighting its impact on EC advancement.
The ablation of PVT1 inhibited the TGF-β/Smad pathway
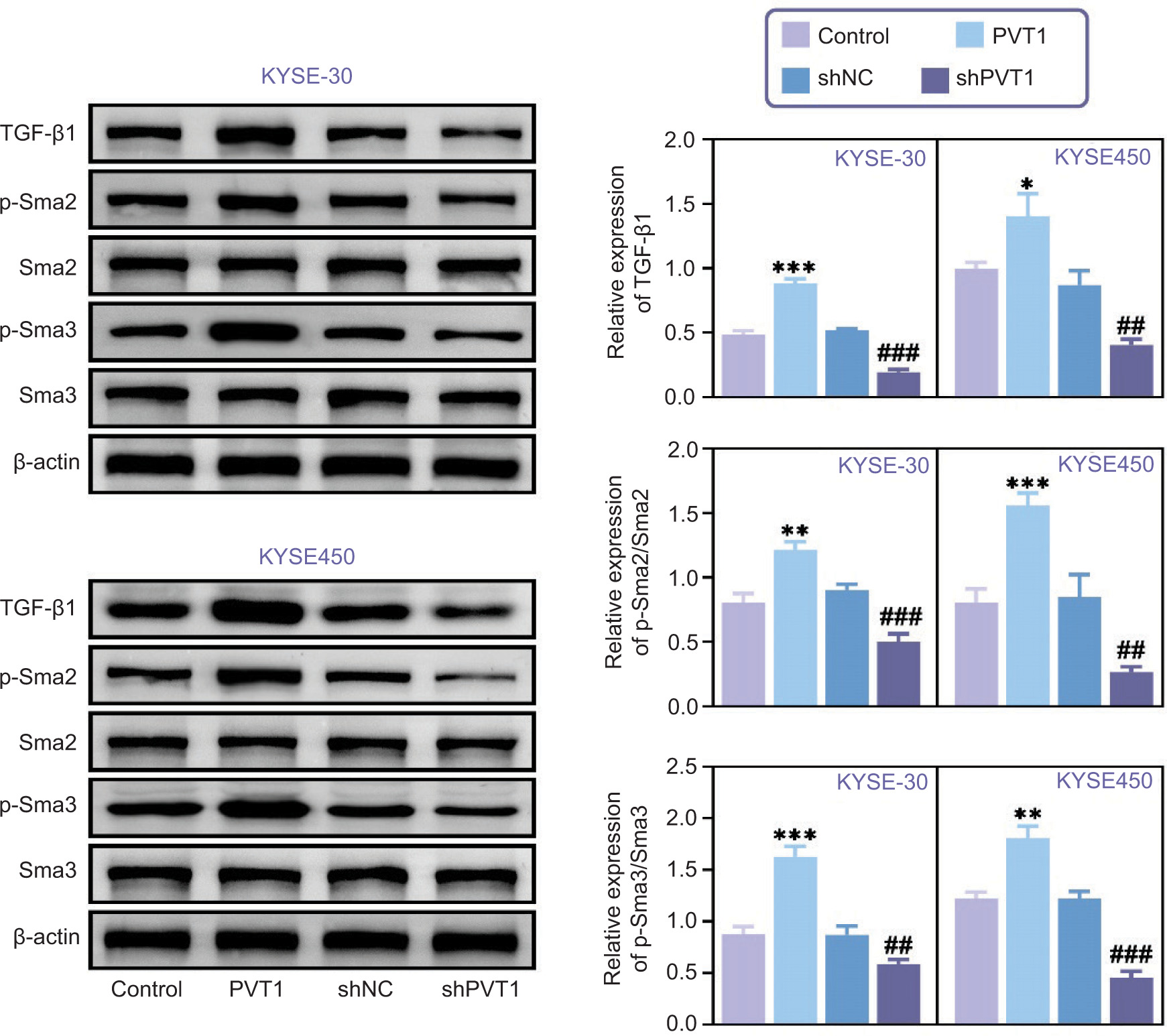
Immunoblot tests have provided us with insights into how PVT1 affects the TGF-β/Smad pathway in KYSE-30 and KYSE-450 cells. When PVT1 levels were reduced, there was a decrease in TGF-β1 expression and the phosphorylation of Smad2 and Smad3, as shown in Figure 3. This decrease implies that the TGF-β/Smad pathway is being suppressed, emphasizing the role of PVT1 in activating this pathway within EC cells. Conversely, overexpression of PVT1 led to an increase in TGF-β1 expression and the phosphorylation of Smad2 and Smad3 in KYSE-30 and KYSE-450 cells, as shown in Figure 3. These results further highlight how PVT1 influences the TGF-β/Smad pathway, confirming its role in activating pathways in cancer cells. Overall, our research shows how PVT1 regulates the dynamics of the TGF-β/Smad pathway in EC cells, highlighting its potential as a target for intervention to combat cancer progression.
Figure 3. The ablation of PVT1 inhibited TGF-β/Smad pathway. Immunoblot assays showed the expression of TGF-β1, Sma2, p-Sma2, Sma3, p-Sma3 in KYSE-30 and KYSE-450 cells upon the indicated transfection. The relative expression was quantified. Error bars indicate SD. *p<0.05, **p<0.01, ***p<0.001, PVT1 versus Control, ##p<0.01, ###p<0.001, shPVT1 versus shNC.
KLT inhibited PVT1/TGF-β/Smad pathway
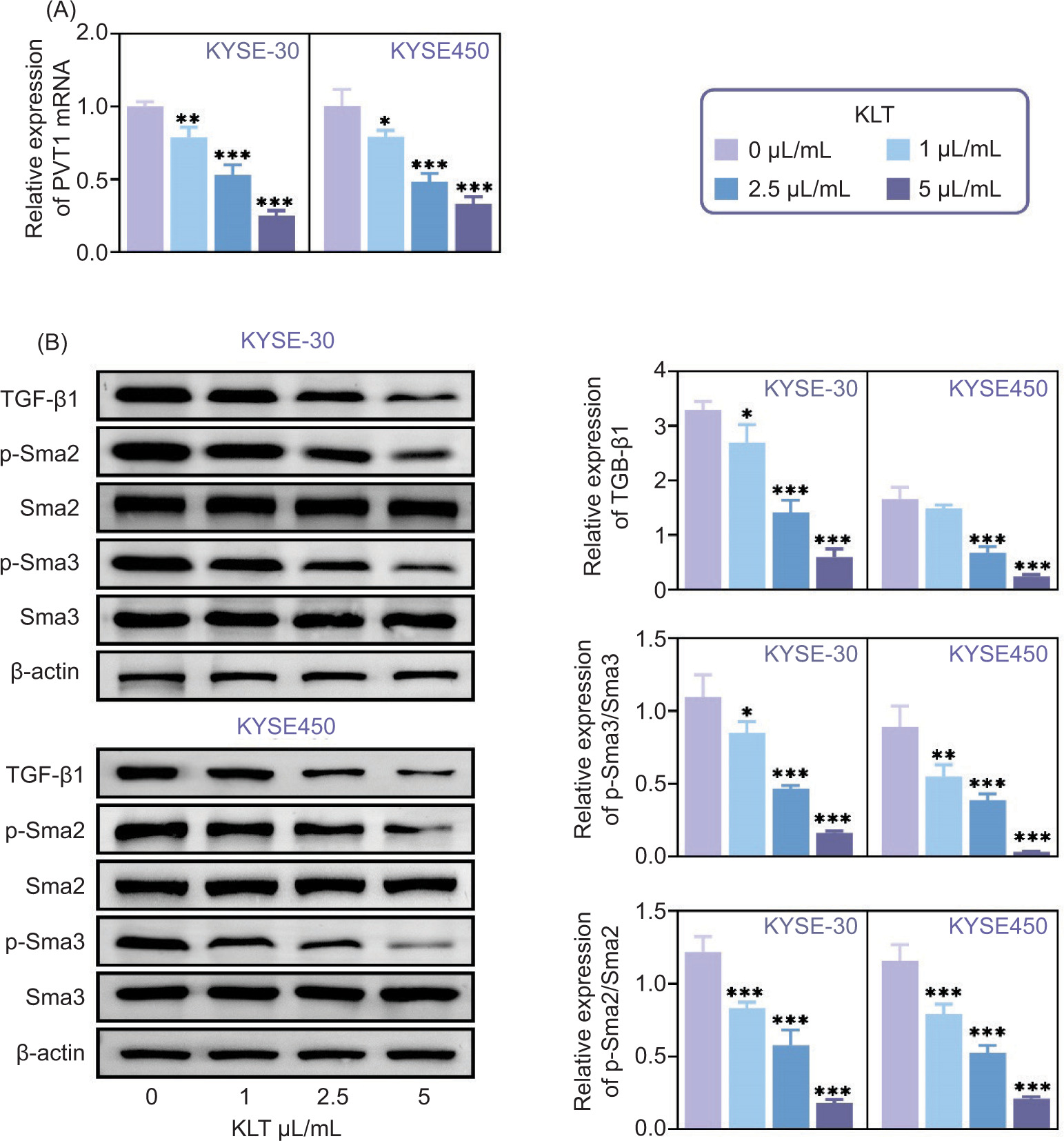
Subsequently, we investigated how KLT affects PVT1 and PVT1/TGF-β/Smad pathways in EC cells. Using qPCR assays, we observed that treating KYSE-30 and KYSE-450 cells with KLT suppressed the mRNA levels of PVT1 in a dose-dependent manner (Figure 4A). Further, immunoblot assays confirmed that increasing doses of KLT reduced the expression of TGF-β1 and the phosphorylation of Sma2 and Sma3 in KYSE-30 and KYSE-450 cells (Figure 4B). This suggests that KLT inhibits the PVT1/TGF-β/Smad pathway in EC cells.
Figure 4. KLT inhibited the PVT1/ TGF-β/Smad pathway. (A) qPCR assays showed the mRNA levels of PVT1 in KYSE-30 and KYSE-450 cells upon the treatment of KLT at the concentrations of 1, 2.5, and 5 µL/mL. (B) Immunoblot assays showed the expression of TGF-β1, Sma2, p-Sma2, Sma3, and p-Sma3 in KYSE-30 and KYSE-450 cells upon the treatment of KLT at the concentrations of 1, 2.5, and 5 µL/mL. Error bars indicate SD. *p<0.05, ***p<0.001.
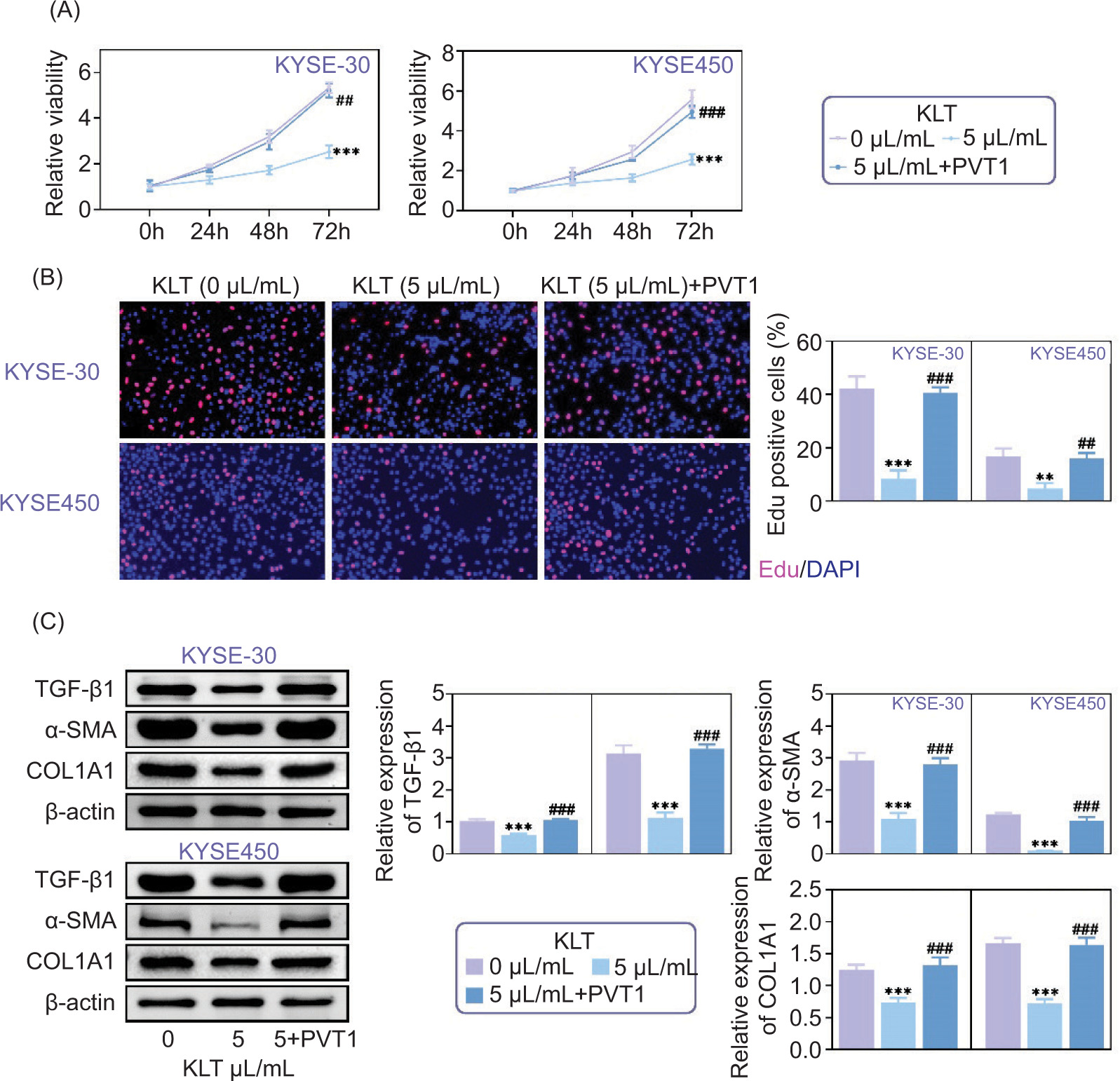
KLT played an antifibrotic role and restrained EC progression by PVT1
Through our research into how Kanglaite (KLT) affects the progression of cancer (EC), we investigated its interaction with PVT1. Using CCK-8 assays, we found that KLT treatment significantly reduced cell viability in KYSE-30 and KYSE-450 cells. Interestingly, when PVT1 was overexpressed, the decrease in cell viability caused by KLT was reversed, as shown in Figure 5A.
Figure 5. KLT played an antifibrotic role and restrained EC progression by PVT1. (A) CCK-8 assays showed the relative viability of KYSE-30 and KYSE-450 cells upon the indicated treatment. The OD450 value was quantified. (B) BrdU assays showed the viability of KYSE-30 and KYSE-450 cells upon the indicated treatment. The BrdU-positive cells were quantified. (C) Immunoblot assays showed the expression of TGF-β1, α-SMA, and COl1A1 in KYSE-30 and KYSE-450 cells upon the indicated treatment. The relative expression was quantified. Error bars indicate SD. ***p<0.001, KLT versus Control, ##p<0.01, ###p<0.001, KLT+PVT1 versus KLT.
Further exploration using EdU assays revealed a pattern indicating that KLT treatment resulted in decrease in the number of EdU cells in KYSE-30 and KYSE-450 cells. Interestingly, the overexpression of PVT1 reversed this decrease, resulting in cell proliferation even in the presence of KLT treatment, as illustrated in Figure 5B.
Immunoblot assays were conducted to uncover the mechanisms behind these observations. They revealed that overexpression of PVT1 restored the reduced expression of TGF-β1, SMA, and COl1A1 induced by KLT treatment, as depicted in Figure 5C. These results indicate that KLT exerts its fibrotic effects and inhibits EC progression by modulating the pathway mediated by PVT1.
Our study emphasizes the relationship between KLT and PVT1 in controlling EC progression. This sheds light on approaches targeting this interaction for managing esophageal cancer.
Discussion
In treating cancer, it is essential to take a personalized approach. Each patient’s unique condition is a key factor in determining the treatment options, including targeted therapy. This personalized method ensures that the treatment plans are customized to address individual needs and characteristics, thereby increasing the likelihood of outcomes. By customizing treatment approaches to fit each patient’s condition, healthcare professionals can effectively manage the complexities of cancer care. From targeted therapies to methods like chemotherapy and surgery, the approach to cancer is constantly evolving, offering hope for improved patient outcomes and quality of life (Lian et al., 2023; Luan et al., 2021a, b; Sun et al., 2023). Esophageal cancer is one of the most common gastrointestinal tumors in clinical practice (Sun et al., 2023). When dealing with cancer, healthcare professionals rely on a combination of three treatments: radiotherapy, chemotherapy, and surgery. These fundamental methods form the basis of cancer management by providing strategies to address the complexities of the disease.
Radiotherapy uses high-energy radiation to target and eliminate cancer cells, either on its own or in combination with other treatments. Chemotherapy involves medications that hinder the growth and spread of cancer cells, often given before or after surgery to enhance results or as a way to ease symptoms in different stages of cancer. On the contrary, surgery entails removing tissue to either cure stage esophageal cancer or provide relief from symptoms and extend survival in more advanced cases.
Together, these treatment options serve as weapons in the fight against cancer, providing patients and healthcare providers with the necessary resources to improve outcomes (Lian et al., 2023). Over the years, there have been advancements in the field of cancer care, providing new hope and opportunities for individuals diagnosed with esophageal cancer. Innovations in treatment options, such as targeted therapies and immunotherapy, have expanded the range of tools to fight this illness.
Despite these advancements, cancer survival rates remain disappointingly low, highlighting the aggressive nature of the disease and the urgent need for innovative research and solutions to improve patient outcomes. While there has been progress, there is still a lot of work to be done to improve early detection methods, develop more efficient treatments, and ultimately increase survival rates for those with esophageal cancer (Yang et al., 2023). In cancer treatment, decisions are carefully customized to fit each patient’s unique situation. For individuals with stage cancer, surgery is often recommended as the primary treatment option. Surgical removal of the tumor provides a chance for a cure and long-term survival.
However, not all patients are candidates for surgery due to factors such as age, overall health condition, or the stage of the disease. In some instances, a combination of radiation therapy and chemotherapy may be advised. This holistic approach, known as chemoradiotherapy, aims to provide treatment by using both radiation and anti-cancer medications to target the tumor.
For individuals with advanced-stage cancer, the focus shifts from curative to palliative care. Palliative care aims to alleviate symptoms, enhance quality of life, and extend survival. Treatment options may include chemotherapy, radiation therapy, targeted therapy, and the emerging field of immunotherapy. Immunotherapy has shown promising advancements in recent years by leveraging the body’s immune system to combat cancer cells.
By tailoring treatment plans to meet the individual needs of each patient, healthcare professionals aim to improve outcomes and support the well-being of those dealing with esophageal cancer (Yang et al., 2020). However, only a few drugs, such as erlotinib, are formulated specifically to target esophageal cancer (Yang et al., 2020). Herein, we have found that KLT plays a role in inhibiting the growth of esophageal cancer and reducing fibrosis by regulating the activity of PVT1/TGF-β/Smad axis. We believe it has the potential to be developed as a therapeutic drug for esophageal cancer.
Our findings reveal the significant role of PVT1 in promoting esophageal cancer (EC) progression and fibrosis by activating the TGF-β/Smad pathway. Specifically, PVT1 was found to be highly expressed in EC tissues and cell lines, correlating with increased expression of fibrosis markers such as TGF-β1, SMA, and COL1A1. Knockdown of PVT1 led to a significant decrease in EC cell growth and fibrosis, as well as reduced activation of the TGF-β/Smad pathway, highlighting its potential as a therapeutic target. Furthermore, treatment with KLT demonstrated its capacity to suppress PVT1 expression and inhibit the PVT1/TGF-β/Smad axis, thereby slowing down EC progression and fibrosis. These results support KLT’s therapeutic promise in managing esophageal cancer by targeting PVT1-mediated pathways.
The results emphasize the importance of PVT1 in promoting the characteristics of EC cells, indicating that it could be a promising target for treating this complex disease. Furthermore, our research highlights the significance of PVT1 by demonstrating that high levels are associated with outcomes in different types of tumors.
By exploring the functions of PVT1 in EC development, our study not only expands our understanding of the condition but also introduces new possibilities for personalized treatments aimed at improving patient prognosis and lifespan (Dong et al., 2023; Li et al., 2022). PVT1 alleviated hypoxia-induced endothelial apoptosis (Zhang et al., 2023). Also, PVT1 inhibited endothelial cell apoptosis in coronary heart disease via mediating MAPK1 expression through targeting miR-532-3p (Liu et al., 2024). Next, we need to detect if PVT1 affects the apoptosis of EC cells. PVT1 enhances gemcitabine resistance in pancreatic cancer by modulating the activation of Wnt/β-catenin and autophagy pathways (Zhang et al., 2023). PVT1 promotes the proliferation and migration of esophageal cancer by regulating the miR-128/ZEB1 axis (Zheng et al., 2016). PVT1 also helps TGF-β/Smad pathway activation, thus promoting tumor progression (Li et al., 2022). Similarly, we also observed a similar phenomenon, and PVT1 also affects EC through the TGF-β/Smad pathway.
Kanglaite (KLT) is a traditional Chinese medicine injection reported to have antitumor activity (Lv et al., 2023; Zhang et al., 2017). KLT has been shown to enhance the efficacy of cisplatin in inhibiting hepatocellular carcinoma by restraining CKLF1-mediated NF-κB pathway (Chen et al., 2021). It has also been found to reverse multidrug resistance of HCC via the PI3K/AKT pathway (Yang et al., 2018) and to suppress EMT caused by TNF-α in colorectal cancer cells (Shi et al., 2018). In addition, KLT has been shown to suppress the expression of drug-resistance proteins by restraining PVT1 in cisplatin-resistant GC cells (Zhang et al., 2017). Furthermore, KLT has been found to regulate the PVT1/TGF-β/Smad axis activity and play an anticancer and antifibrotic role in EC.
Members of the TGF-β family are known to be powerful inducers of EMT in various physiological and pathological conditions (Rasouli et al., 2023). The abnormal expression of TGF-β1 has been well documented in tumor microenvironments and fibrotic lesions (Rasouli et al., 2023). In addition, tumor fibrosis can affect drug delivery, hinder the accessibility of immune cells, and promote disease invasion and treatment resistance. Therefore, inhibiting the occurrence of tumor fibrosis is necessary (Tang et al., 2022). Our research indicates that PVT1 promotes the growth and scarring of cancer by triggering the TGF-β/Smad pathway. KLT shows promise for treating both cancer and fibrosis in esophageal cancer cases.
In summary, Kanglaite (KLT) shows abilities in fighting tumors and fibrosis in cancer (EC) by effectively blocking the PVT1/TGF-β/Smad pathway. These findings highlight KLT’s potential as a medication for treating EC.
Acknowledgments
Not applicable.
Funding
This study is funded by Zhenjiang City Science and Technology Innovation Fund (Key Research and Development Plan – Social Development) Project (Grant No. SH2023060) and Open Project of Jiangsu Key Laboratory of Basic and Clinical Transformation of Non-coding RNA (Grant No. 202110).
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
The datasets used and/or analyzed during the present study are available from the corresponding author upon reasonable request.
Conflicts of Interest
The authors state that there are no conflicts of interest to disclose.
Ethics Approval
Ethical approval was obtained from the Ethics Committee of the Third People’s Hospital of Zhenjiang.
Statement of Informed Consent
Written informed consent was obtained from a legally authorized representative(s) for anonymized patient information to be published in this article.
Contribution of Authors
Xiuqing Li and Ka Bian designed the study, completed the experiment, and supervised the data collection; Ji Zhang analyzed and interpreted the data; Kaili Luo, Tao Dong, and Chenglin Luo prepared the manuscript for publication and reviewed the draft of the manuscript. All authors have read and approved the manuscript.
REFERENCES
Chen, C., Ai, Q. D., & Wei, Y. H. (2021). Kanglaite enhances the efficacy of cisplatin in suppression of hepatocellular carcinoma via inhibiting CKLF1 mediated NF-kappaB pathway and regulating transporter mediated drug efflux. Journal of Ethnopharmacology, 264, 113388. 10.1016/j.jep.2020.113388
Dong, L., Wang, H., Gao, Y., Wang, S., & Wang, W. (2023). Long non-coding RNA PVT1 promotes the proliferation, migration and EMT process of ovarian cancer cells by regulating CTGF. Oncology Letters, 25(2), 71. 10.3892/ol.2022.13657
Kashiwagi, M., Ojima, T., Hayata, K., Kitadani, J., Takeuchi, A., Kuroi, A., et al. (2023). Impact of pulmonary vein anatomy and left atrial size on postoperative atrial fibrillation after esophagectomy for esophageal cancer. Esophagus, 20(4), 626–634. 10.1007/s10388-023-01018-8
Li, Z., Li, M., Xia, P., Wang, L., & Lu, Z. (2022). Targeting long non-coding RNA PVT1/TGF-beta/Smad by p53 prevents glioma progression. Cancer Biology & Therapy, 23(1), 225–233. 10.1080/15384047.2022.2042160
Lian, J., Xu, A., Chen, T., Zhang, H., & Xu, M. (2023). Unexpected gastric perforation during endoscopic submucosal tunnel dissection for early circumferential esophageal cancer. Endoscopy, 55(S 01), E833–E834. 10.1055/a-2106-1012
Liu, H., Ma, X. F., Dong, N., Wang, G. N., Qi, M. X., & Tan, J. K. (2024). LncRNA PVT1 inhibits endothelial cells apoptosis in coronary heart disease through regulating MAPK1 expression via miR-532-3p. Acta Cardiologica, 79(3), 295–303. 10.1080/00015385.2023.2209448
Liu, L., Huan, L., Zhang, Y., Wei, W., Chen, Z., Xu, D., et al. (2022). Ubiquitin-specific protease 8 inhibits lipopolysaccharide-triggered pyroptosis of human bronchial epithelial cells by regulating PI3K/AKT and NF-kappaB pathways. Allergol Immunopathol (Madr), 50(2), 96–103. 10.15586/aei.v50i2.568
Luan, Y., Luan, Y., Feng, Q., Chen, X., Ren, K. D., & Yang, Y. (2021a). Emerging role of mitophagy in the heart: Therapeutic potentials to modulate mitophagy in cardiac diseases. Oxidative Medicine and Cellular Longevity, 2021, 3259963. 10.1155/2021/3259963
Luan, Y., Ren, K. D., Luan, Y., Chen, X., & Yang, Y. (2021b). Mitochondrial dynamics: Pathogenesis and therapeutic targets of vascular diseases. Frontiers in Cardiovascular Medicine, 8, 770574. 10.3389/fcvm.2021.770574
Luo, Z., Chen, R., Hu, S., Huang, X., & Huang, Z. (2022). PVT1 promotes resistance to 5-FU in colon cancer via the miR-486-5p/CDK4 axis. Oncology Letters, 24(2), 280. 10.3892/ol.2022.13400
Lv, P., & Liu, M. (2023). Meta-analysis of the clinical effect of Kanglaite injection-assisted gemcitabine plus cisplatin regimen on non-small cell lung cancer. American Journal of Translational Research, 15(5), 2999–3012.
Rasouli, S., Heshmatnia, J., Mosaffa, N., Marjani, M., & Mortaz, E. (2023). Dysregulation of immunity in pulmonary fibrosis is associated with increased myeloid-specific triggering receptor-1 and transforming growth factor-beta1 expression. Iranian Journal of Allergy, Asthma, and Immunology, 22(1), 12–24. 10.18502/ijaai.v22i1.12002
Shi, G., Zheng, X., Zhang, S., Wu, X., Yu, F., Wang, Y., et al. (2018). Kanglaite inhibits EMT caused by TNF-alpha via NF-kappaBeta inhibition in colorectal cancer cells. Oncotarget, 9(6), 6771–6779. 10.18632/oncotarget.23645
Sun, C., Ye, Y., Tan, Z., Liu, Y., Li, Y., Hu, W., et al. (2023). Tumor-associated nonmyelinating Schwann cell-expressed PVT1 promotes pancreatic cancer kynurenine pathway and tumor immune exclusion. Science Advances, 9(5), eadd6995. 10.1126/sciadv.add6995
Sun, J., Sun, Y., Miniderima, & Wang, X. (2023). Cytokine-induced killer cell treatment is superior to chemotherapy alone in esophageal cancer. Pathology Oncology Research, 29, 1610710. 10.3389/pore.2023.1610710
Tang, P. C., Zhang, Y. Y., Li, J. S., Chan, M. K., Chen, J., Tang, Y., et al. (2022). LncRNA-dependent mechanisms of transforming growth factor-beta: From tissue fibrosis to cancer progression. Non-coding RNA, 8(3), 26. 10.3390/ncrna8030036
Wang, P., Chen, C., Zhang, Q. H., Sun, G. D., Wang, C. A., & Li, W. (2021). Retraction of lumbar disc herniation achieved by noninvasive techniques: A case report. World Journal of Clinical Cases, 9(27), 8082–8089. 10.12998/wjcc.v9.i27.8082
Wu, B. Q., Jiang, Y., Zhu, F., Sun, D. L., & He, X. Z. (2017). Long noncoding RNA PVT1 promotes EMT and cell proliferation and migration through downregulating p21 in pancreatic cancer cells. Technology in Cancer Research & Treatment, 16(6), 819–827. 10.1177/1533034617700559
Yanagida, M., Hosoi, Y., Kawano, T., Otake, Y., Yamanaka, Y., Baba, T., et al. (2024). Noniatrogenic meningitis caused by Streptococcus salivarius associated with early esophageal cancer and early gastric cancer. Internal Medicine, 63(3), 457–460. 10.2169/internalmedicine.1304-22
Yang, C., Hou, A., Yu, C., Dai, L., Wang, W., Zhang, K., et al. (2018). Kanglaite reverses multidrug resistance of HCC by inducing apoptosis and cell cycle arrest via PI3K/AKT pathway. OncoTargets and Therapy, 119, 83–96. 10.2147/OTT.S153814
Yang, X. X., Ma, M., Sang, M. X., Zhang, X. Y., Zou, N. Y., & Zhu, S. C. (2020). Knockdown of FAM83D enhances radiosensitivity in coordination with irradiation by inhibiting EMT via the Akt/GSK-3beta/Snail signaling pathway in human esophageal cancer cells. OncoTargets and Therapy, 13, 4665–4678. 10.2147/OTT.S245681
Yang, Y. M., Hong, P., Xu, W. W., He, Q. Y., & Li, B. (2020). Advances in targeted therapy for esophageal cancer. Signal Transduction and Targeted Therapy, 5(1), 229. 10.1038/s41392-020-00323-3
Zhang, P., Gong, S., Li, S., & Yuan, Z. (2023). PVT1 alleviates hypoxia-induced endothelial apoptosis by enhancing autophagy via the miR-15b-5p/ATG14 and miR-424-5p/ATG14 axis. Biochemical and Biophysical Research Communications, 67, 11–19. 10.1016/j.bbrc.2023.06.001
Zhang, X. W., Liu, L., Zhang, X. Z., & Bo, P. (2017). Kanglaite inhibits the expression of drug resistance genes through suppressing PVT1 in cisplatin-resistant gastric cancer cells. Experimental and Therapeutic Medicine, 14(2), 1789–1794. 10.3892/etm.2017.4650
Zhao, M., Fu, L., Xu, P., Wang, T., & Li, P. (2023). Network pharmacology and experimental validation to explore the effect and mechanism of Kanglaite injection against triple-negative breast cancer. Drug Design, Development and Therapy, 17, 901–917. 10.2147/DDDT.S397969
Zheng, X., Hu, H., & Li, S. (2016). High expression of lncRNA PVT1 promotes invasion by inducing epithelial-to-mesenchymal transition in esophageal cancer. Oncology Letters, 12(4), 2357–2362. 10.3892/ol.2016.5026