
Download
PAPER
Study of oxygen in wines with different proportions of yeast lees
Michaela Kulhankova, Bozena Prusova*, Mojmir Baron
Department of Viticulture and Enology, Mendel University in Brno, Valticka, Lednice, Czech Republic
Abstract
The lees produced during fermentation are known for their ability to consume oxygen. During wine aging, it has a positive effect on the antioxidant and sensory properties of wine. This study focuses on the effects of different doses of fermentation lees on the oxygen consumption and antioxidant activity of wine, which are important for the quality of the final product as well as consumer. The effect on oxygen consumption after bottling, antioxidant activity and phenolic composition was studied for wines of the variety Grüner Veltliner with different proportions of yeast lees. The rate of dissolved oxygen consumption increased with increasing dosages of fermentation lees. The first significant decrease was observed as early as the second day after bottling for all variants above 20 g of fermentation lees in 0.75 L of wine. Total polyphenol content and antioxidant activity also increased with the dose of yeast lees. The major negative parameter for the highest addition of lees was the concentration of volatile acids, which increased from a baseline value of 0.34 g L–1 to 0.45 g L–1.
Key words: antioxidant activity, oxygen, phenolic compounds, wine oxidation, yeast lees
*Corresponding Author: Bozena Prusova, Department of Viticulture and Enology, Mendel University in Brno, Valticka 337, 691 44 Lednice, Czech Republic. Email: [email protected]
Received: 12 December 2022; Accepted: 25 April 2023; Published: 5 January 2024
© 2024 Codon Publications
This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 International (CC BY-NC-SA 4.0). License (http://creativecommons.org/licenses/by-nc-sa/4.0/)
Introduction
Oxygen in wine plays a crucial role from a chemical and biological standpoint. It affects the wine through grape harvesting, processing, fermentation and maturation, whether stored in steel tanks, wooden barrels or bottles. Finding a balance between oxidation and reduction is a complex process in which it is necessary to understand the reactions of oxygen with the substances it reacts. Oxygen management using fermentation lees has a positive effect on the quality of the resulting wine and its antioxidant properties, and thus minimises the dosage of sulphur dioxide (Baron and Sochor, 2013; Day et al., 2015; Vidal and Moutounet, 2008; Zironi et al., 2010).
Oxygen affects wine during two phases of the winemaking process: the dynamic process phases (i.e. bottling, filtration and mixing), and the static phases (i.e. during storage in tanks, barrels and bottles). Oxygen diffuses through holes in containers (bottle neck) or through the material itself (e.g. wood, seal or cork) (Valade et al., 2006). The wine is affected either negatively or positively by quantity, timing and its chemical composition and physical properties, especially temperature. From a chemical perspective, wine primarily affects the modification of phenolic compounds. In case of red wines, it can favourably affect the colour of the wine because of polymerisation. In white wines, it may cause browning and change in colour because of the oxidation of polyphenols. However, it has positive effects on the development and maturation of wine from a sensory point of view. Furthermore, oxygen affects aroma and multiplication and growth of microorganisms (Eisenman, 1987; Zironi et al., 2010).
An important part of the technology is the use of yeast lees. Suspended lees have a high capacity of oxygen consumption even several years after the end of fermentation, thus protecting the wine against oxidation immediately after fermentation as well as throughout the entire maturation process. Perez-Serradilla and de Castro (2008) place the use of yeast lees against oxidation among traditional wine protection systems. This claim is confirmed by Schneider et al. (2016) that the high antioxidant properties of lees are mainly caused by yeast lipids and sterols.
Oxygen consumption by yeast lees is affected by various technological processes (e.g. filling barrels, regular stirring of the lees, or refilling containers). All these furnish variable degree of support in the dissolution of oxygen in wine (Fornairon-Bonnefond and Salmon, 2003).
Schneider et al. (2016) conducted a similar study. Fermentation was carried out using six types of commercial active dry yeasts in both white and red wines. Yeast dosage was performed via turbidity (nephelometric turbidity unit [NTU]) measurements using a pre-established regression between yeast cell numbers and NTU. This study includes an experiment with one yeast strain but finds the ideal in yeast lees dosage.
The aim of this study was to better understand the effect of fermentation lees on the antioxidant properties of wine in relation to oxygen management and consequently the possibility of reducing the doses of sulphur dioxide. The novelty of the study existed in the more detailed investigation of the effect of different proportions of fermentation lees on the antioxidant potential and chemical composition of wine. The wine variety Grüner Veltliner was chosen for its neutrality and flexibility. Thanks to its low aromatic intensity, this variety could be used for both light and fresh wines and wines of a more complex structure using lees.
Material and Methods
Design of experiment
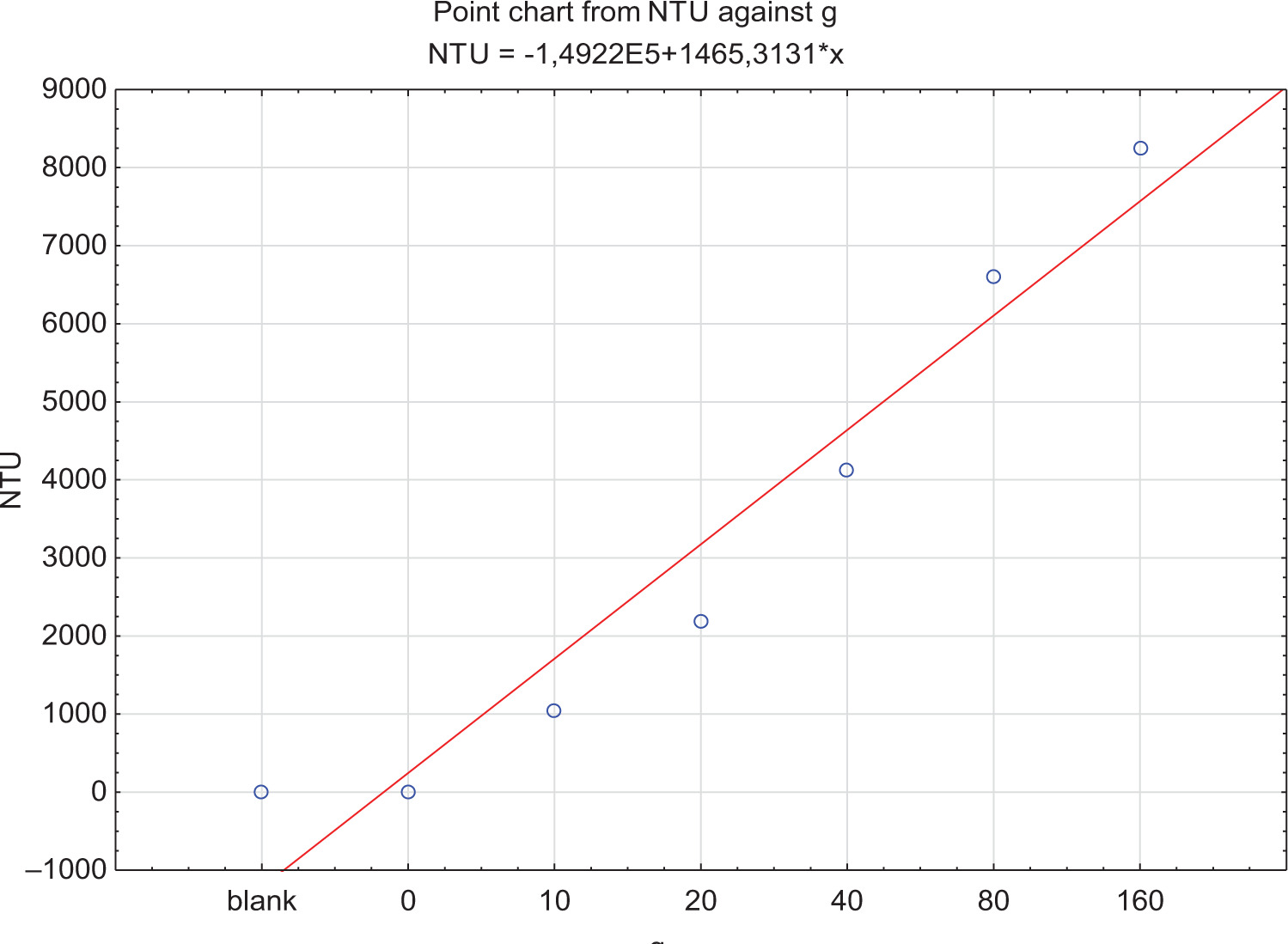
The study obtained clarified Grüner Veltliner wine variety from the regular and conventional production of 2020, and separated yeast lees produced during fermentation of the sample wine. The grapes for wine production were harvested from Dolni Dunajovice, a village at Mikulov in the wine region of Moravia (Czech Republic). The vineyard has loamy soil and is located on a southwest-facing slope. The analytic parameters of the musth were: pH 3.2, sugar content 22.6 Brix, total acidity 8.9 g L–1, and assimilable nitrogen 225 mg L–1. The musth was inoculated with a dose of 15 g hL–1 of the commercial active dry yeast Oenoferm Klosterneuburg (Erbslöh, Geisenheim, Germany). After fermentation, the wine was decanted from fermentation lees, which were stored in a separate container; then the wine was clarified with a dose of 80 g hL–1 of Bentostab (IOC, Épernay, France). The wine was divided into six 0.75-L variants in three repetitions with doses of 0 g (control sample/8.78 NTU), 10 g (1.34 wt.%/1047.67 NTU), 20 g (2.68 wt.%/2189.67 NTU), 40 g (5.37 wt.%/4,770 NTU), 80 g (10.74 wt.%/6,600.67 NTU), and 160 g (21.47 wt.%/8256.33 NTU) of post-fermentation lees primarily containing yeast biomass (hereinafter referred to as yeast lees) (see Figure 1); from this, 50-mL samples were taken and frozen for analysis. The lees were added in wet form to obtain the required NTU values with a linear curve. This method of lees dosing is a practical method for subsequent replication in a winery. Experiments conducted in 750-mL bottles described more accurately the oxygen consumption model directly in the bottles, giving us a better insight into the development of oxygen concentration, which could be later extrapolated to non-filtered wines. This method of measurement is more suitable for longer-term monitoring and minimises environmental effects.
Figure 1. Correlation between turbidity (NTU) and lees content (g) in a 750-mL bottle.
NomaSense O2 P6000 (manufacturer Nomacorc, Italy), a portable oxygen analyser, was applied to 750-mL bottles for monitoring headspace gaseous oxygen levels in two areas of the bottles: the bottle neck for monitoring dissolved oxygen (DO) in the headspace, and the lower one-third of the bottle for monitoring dissolved oxygen in the wine. A predetermined quantity of yeast lees was transferred to the bottles, after which the bottles were filled with wine to a standard level using the semi-automatic vacuum filler of Enolmatic bottles (manufacturer TENCO, Avegno, Italy) and fitted with a cork closure (DIAM finely milled cork).
NomaSense oxygen measurement
NomaSense portable analyser (Nomacorc) was used to measure oxygen level in wine by applying luminescence technology combined with remote sensors and other accessories. This analyser can measure dissolved oxygen in both headspace and wine. The sensor has a built-in barometer and temperature sensor to measure dissolved oxygen directly in the vessel, allowing optimal measurement accuracy for concentrations ranging from 0 mg L–1 to 22 mg L–1 with an accuracy of ±0.04 mg L–1 and a detection limit of 15 μg L–1 (Vinventions, 2018; Winebussines, 2019).
WineScan to measure basic wine parameters
The WineScanTM Auto (manufacturer FOSS, Denmark) analyser was used to measure basic analytical parameters of the wine. It works on the principle of Fourier transform infrared spectroscopy (FTIR). It uses the entire infrared spectrum and simultaneously analyses various parameters with high accuracy. Each analysis was conducted for three times. The interferometer of this apparatus was fully encapsulated to minimise any interference by water vapour and other gases (e.g. CO2) in the optical path (Friedel et al., 2013).
Determination of total phenols
Total phenols were determined using the modified Folin–Ciocalteu method. Samples of 12-μL and 10-μL Folin–Ciocalteu reagent were added to 198 μL of wine. After 36 s, 30 μL of 20% sodium carbonate solution was added. Absorbance at 700 nm was measured after 600 s. The concentration of total phenols was calculated on a calibration curve using gallic acid equivalent (GAE) as the standard (25–1,000 mg L–1). The results were expressed as GAE (Sochor et al., 2014; Sochorova et al., 2020; Waterman and Mole, 1994).
Determination of antioxidant activity by 2,2-Diphenyl-1-picrylhydrazyl (DPPH) method
Antioxidant activity was determined by the method previously described by Sochor et al. (2010). A sample of 150 μL of (0.095-mM DPPH reagent was incubated in 15 μL of wine. The absorbance was measured for 10 min at 505 nm, and the output ratio was calculated as a difference between the absorbance values measured in the 10th minute and 2nd minute of the test (Carmona-Jimenez et al., 2014; Sochor et al., 2014). Antioxidant activity was determined in three repetitions immediately after sampling.
Determination of antioxidant activity by ferric reducing antioxidant power (FRAP) assay
A 12-μL wine sample was added to 198 μL of alkaline buffer containing 200-mM sodium acetate and treated with acetic acid to a pH of 3.6, then 20 μL of 20-mM FeCl3 and 20-μL 10-mM 2,4,6-tripyridyl-s-triazine solution dissolved 40-mM HCl were added. The reduction force was calculated from the calibration curve using ascorbic acid (0.1–3 mM) or gallic acid (10–300 mg L–1) as a standard. Results were expressed in GAE (Pulido et al., 2000).
Gas chromatography–mass spectrometry (GC-MS) analysis aromatic profile
The concentration of individual volatile substances in the wine was determined according to the methyl tert-butyl ether extraction (MTBE) method. A 20-mL wine sample was pipetted into a 25-mL volumetric flask containing 50-μL 2-nonanol solution in ethanol as an internal standard (400 mg.L–1) and 5-mL saturated solution of (NH4)2SO4 (Prusova and Baron, 2018). Solution in the flask was thoroughly mixed and subsequently added to 0.75 mL of extraction solvent (MTBE with addition of 1% cyclohexane). After thorough mixing, the individual phases were separated, and the upper organic layer and the emulsion were transferred into a microtube. The microtube content was centrifuged and the upper clear proportion was dried with anhydrous magnesium sulphate prior to GC-MS analysis. Extraction and subsequent GC-MS analysis were performed in three repetitions. Average of the resulting values was calculated, and the standard deviation was determined using Excel and Statistic 10.
The experiment was performed in a GC Shimadzu (GC-17A) equipped with an AOC-5000 autosampler and connected to a quasi-peak detector (QP-5050A). Subsequent identification was made using GC-MS solution software (LabSolutions software, version 1.20). Separation conditions for the analysis were as follows: column DB-WAX 30 m × 0.25 mm, stationary phase 0.25 mm (polyethylene glycol), detector voltage 1.5 kV, volume of the sample injected 1 μL, and partition ratio 1:5. The carrier gas was helium (He) with a flow rate of 1 mL min–1 (linear gas speed was 36 cm s–1), and the spray temperature was 180°C. The initial column temperature was 45°C, which was maintained for 3.5 min, followed by the following temperature gradients: up to 75°C, gradient of 6°C.min–1; up to 126°C, gradient of 3°C min–1; up to 190°C, gradient of 4°C min–1; and up to 250°C, gradient of 5°C.min–1. The final temperature was maintained for 6.5 min, with the overall analysis lasting 60 min. The detector was operated in SCAN mode with an interval of 0.25 s in the range of 14–264 s. Individual compounds were identified by comparing GC-MS spectra and retention time spectra with NIST 107.
Results and Discussion
Results
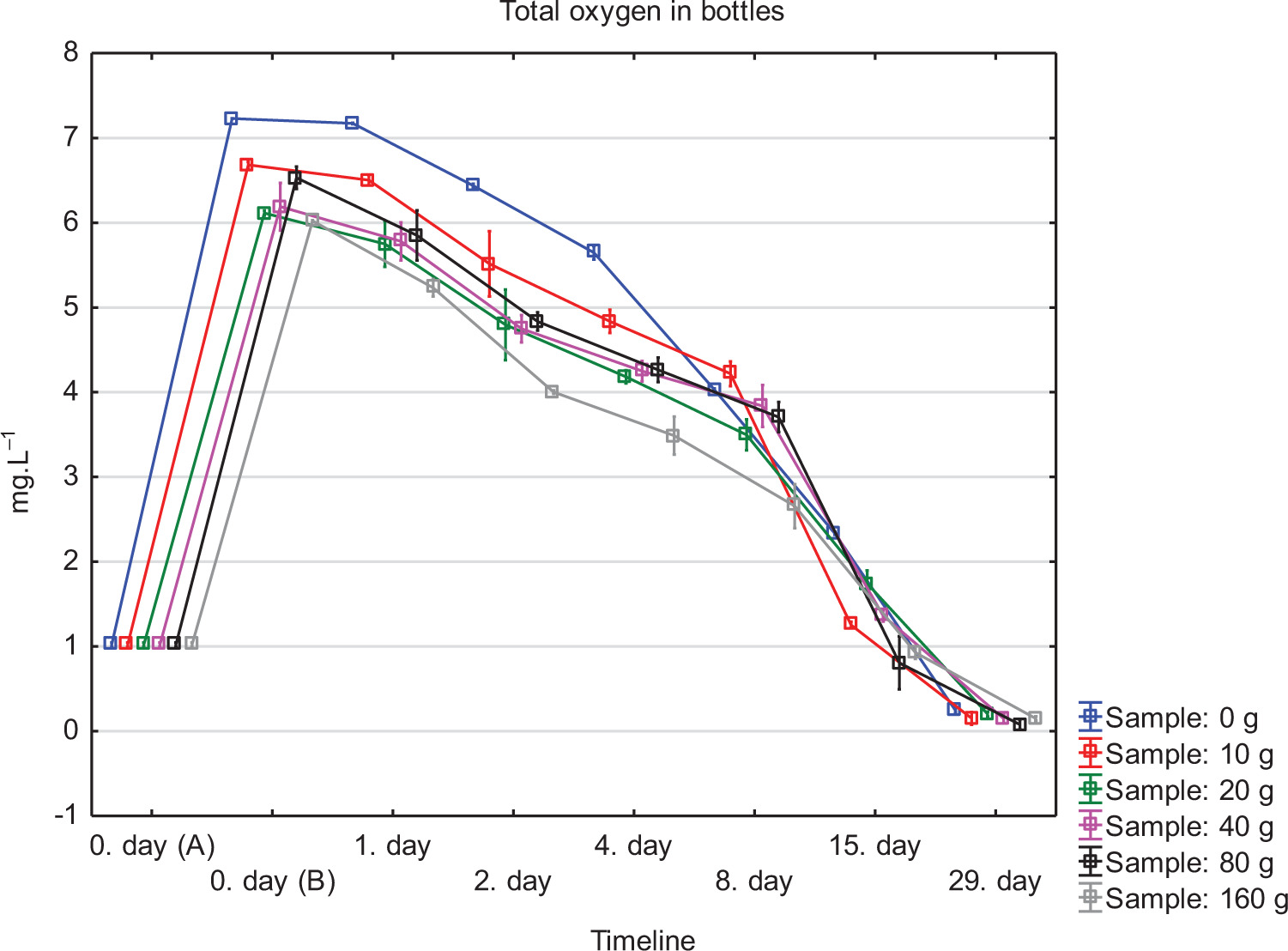
The experiment aimed to evaluate the effect of different doses of yeast lees on dissolved oxygen concentration and chemical composition of bottled wine samples (see Figure 2). The experiment evaluated six variants of 0–160-g yeast lees in 750-mL bottles. The results are shown in Table 1.
Figure 2. Total oxygen in bottles of all sample variants of wine in the timeline.
Table 1. Basic analytical parameters of base wine and its variants treated with yeast lees.
| Sample name | Gluc/Fruct (g L–1) |
pH | Total acids (g L–1) |
Volatile acids (g L–1) |
Malic acid (g L–1) |
Lactic acid (g L–1) |
Glycerol (g L–1) |
|---|---|---|---|---|---|---|---|
| 0-g yeast lees | 3.53±0.06a | 3.14±0.01a | 6.40±0.06b | 0.35±0.01a | 0.80±0.03a | 0.50±0.03c | 5.67±0.06a |
| 10-g yeast lees | 3.60±0.01b | 3.14±0.00a | 6.60±0.01a,b | 0.35±0.03a | 0.83±0.06a,b | 0.63±0.06a | 5.67±0.06a |
| 20-g yeast lees | 3.80±0.03c | 3.15±0.00b | 6.70±0.01a | 0.36±0.01a | 0.83±0.06a,b | 0.70±0.10a,b | 5.70±0.03a |
| 40-g yeast lees | 4.00±0.01d | 3.17±0.01c | 6.60±0.01a | 0.38±0.01b | 0.83±0.06a,b | 0.70±0.03a,b | 5.80±0.03a,b |
| 80-g yeast lees | 4.33±0.03e | 3.20±0.00d | 6.60±0.01a,b | 0.39±0.01c | 0.90±0.03b | 0.80±0.03b | 5.90±0.03b |
| 160-g yeast lees | 4.80±0.03f | 3.30±0.00e | 6.50±0.06a,b | 0.45±0.01d | 0.40±0.03c | 1.43±0.06d | 6.20±0.03c |
Note: Results are expressed as the mean value of three measurements ± standard deviation. The division into homogeneous groups (a,b,c,d,e,f) was based on Fisher’s test, the significance level is α = 0.005.
Basic analytical parameters of the wine were determined prior to the experiment. These were essential for the comparison of individual chemical parameters, depending on the amount of yeast lees added. The most significant changes were discovered in the amount of volatile acids and the lactic acid–malic acid ratio. Changes observed in all these parameters were due to the non-sterility of wine when malolactic fermentation took place. Increase in pH was proportional to the conversion of malic acid to lactic acid during malolactic fermentation.
Figure 2 the dates 0, 1, 2, 4, 8, 15 and 29 from filling using spots placed in two places inside the bottle – head space and inside of wine, as described before.
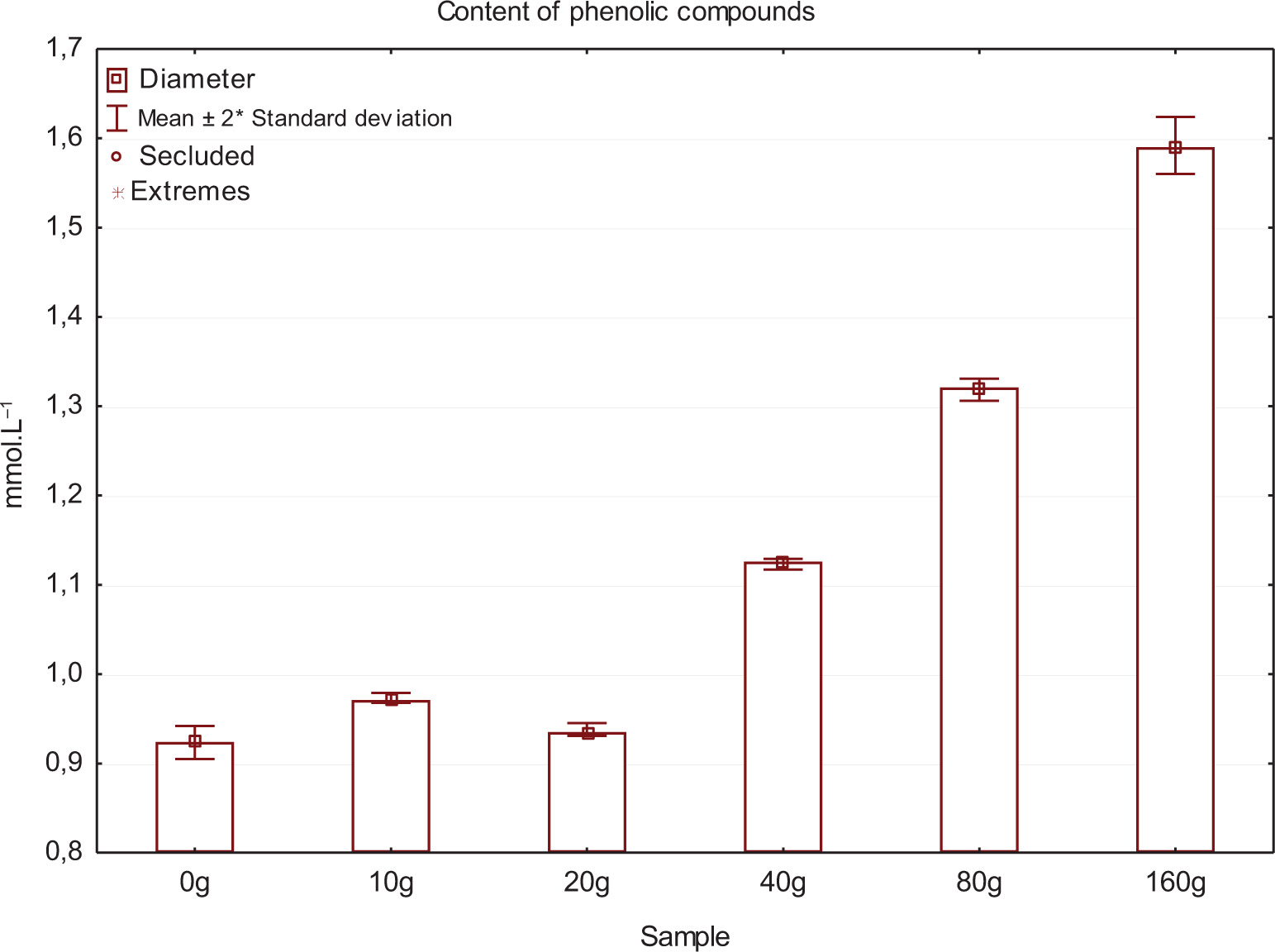
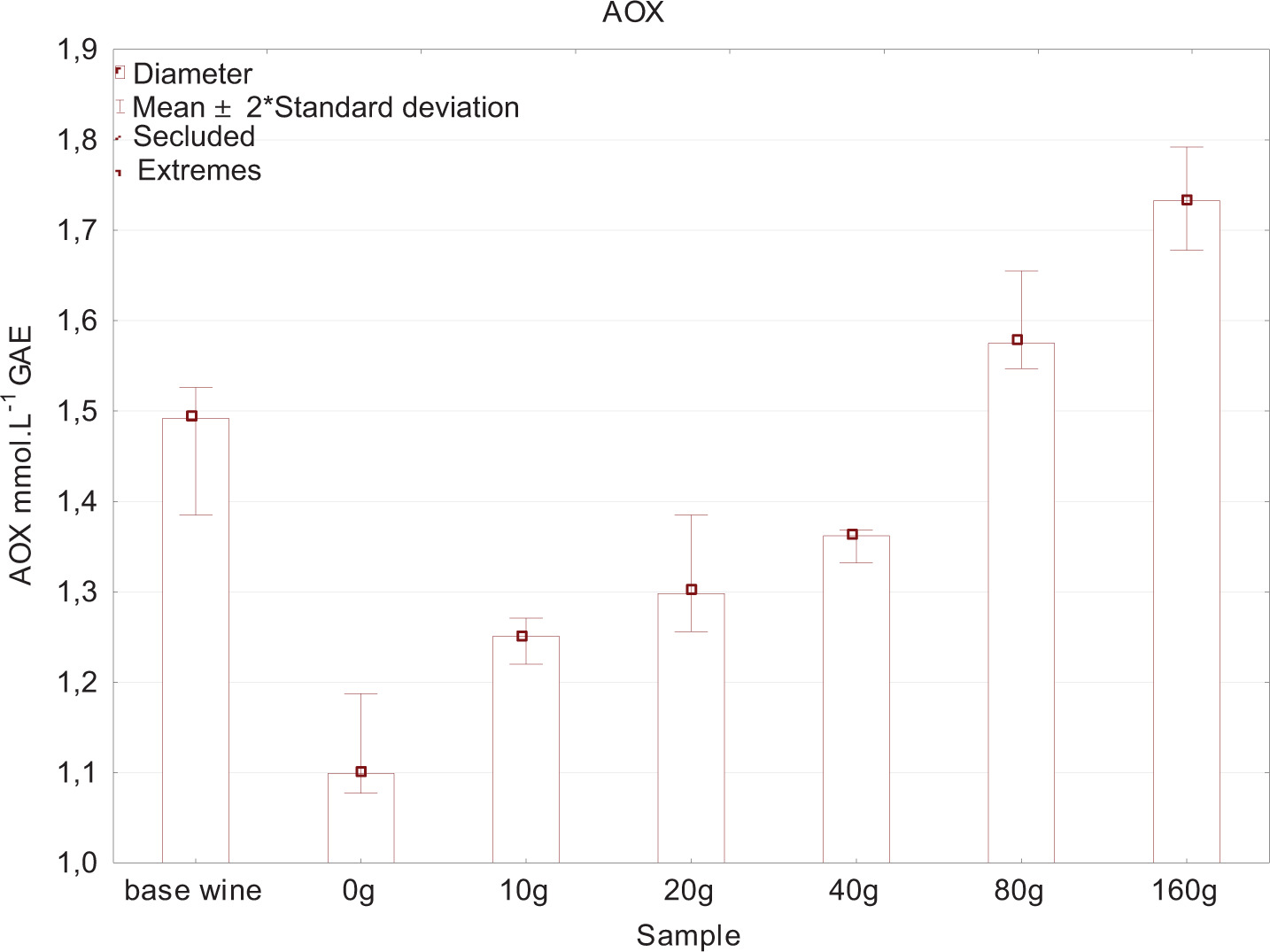
The experiment demonstrated that different proportions of yeast lees contained in the wine affected the rate of dissolved oxygen consumption. In Figure 2, the initial values indicate the concentration of oxygen in its entire volume before the bottles were filled. Further concentrations of oxygen were measured immediately after bottling and then at intervals as shown in the figure. Bottling caused aeration of all samples; this is shown as an increase in curve on day 0. After this day, a decrease in oxygen was observed in individual variants. The fastest drop in concentration was observed in 160-g yeast lees variant, where 0.91 mg L–1 of oxygen was determined when measured on day 15, compared to 2.34 mg L–1 of oxygen for 0 g yeast lees variant measured on the same day. This confirmed the results of the study conducted by Schneider et al. (2016) with six yeast strains (EC 1118, CY 3079, Fermicru VB1 etc.) to assess dissolved oxygen consumption by yeast lees. The cited study observed that the oxygen consumption curve over time initially had a linear increase, followed by a transition to a steady state. This demonstrates the depletion of oxygen sources by yeast lees; after 45–55 h, 40–60 mg L–1 dissolved oxygen was consumed. As a result, even a small amount of suspended lees (equivalent to 50 NTU) was enough to consume the oxygen that has accumulated in a bottle. Schneider et al. (2016) stated that the rate of dissolved oxygen uptake by yeast lees could be between 0.003 mg L–1 and 0.011 mg L–1 per 10–9 cells from the second to sixth month of aging at 14°C. Yeast lees contain a large number of polyphenols that increase antioxidant properties. This explains the presence of significantly higher concentrations of total phenols in the experiment for variants with a higher proportion of yeast lees (Figure 3); polyphenol content increases with increasing doses of fermentation lees. Another explanation would be that yeast cells contain and release into wine the following organic compounds: glutathione, tyrosol and various complexes, including hydroxycinnamates, amino acids with a phenolic -OH group, and peptides containing methionine, tryptophan, and tyrosine, all of which are included in the measurement using Folin–Ciocalteu reagent (Comuzzo et al., 2015; Myers and Singleton, 1979; Singleton and Rossi, 1965).
Figure 3. Total polyphenol content in all wine samples.
In general, phenolic compounds in wine appear from four main sources: berries or other parts of vines, action of microorganisms during fermentation, oak used in the maturation of wine (e.g. barrels or chips), and commercial tannins added during the production process (Zhang et al., 2021).
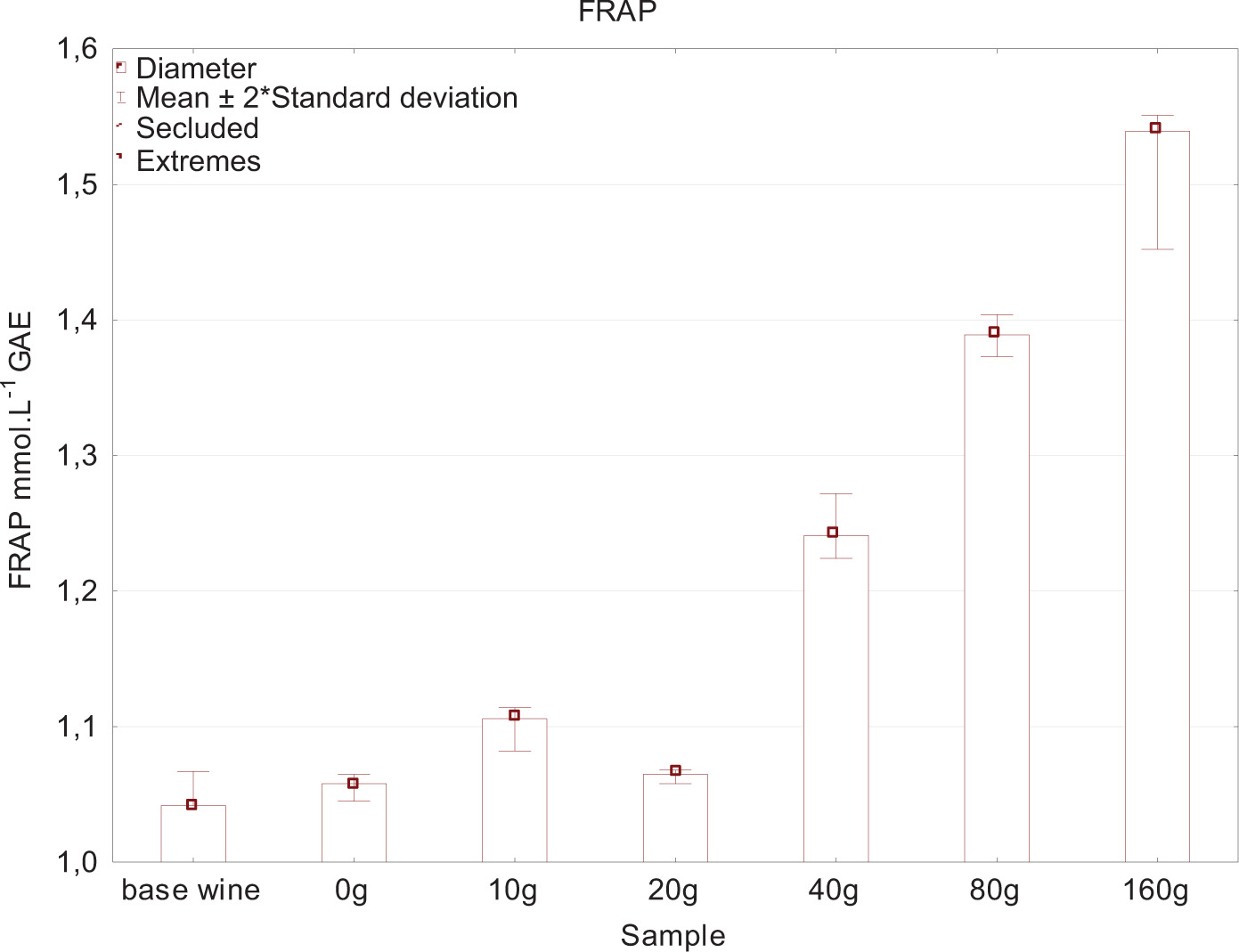
Figures 4 and 5 show that antioxidant activity increases with increasing the addition of yeast lees because of oxygen consumption by both polyphenols and yeast. Salmon et al. (2002) discovered that polyphenol fractions dissolved in wine were more reactive, while polyphenols adsorbed with yeast lees were slightly reactive to oxygen. Laboratory analysis included the determination of aromatic profile of individual wines. For clarity, these results were processed and tabulated statistically (Tables 2 and 3). Comparisons were performed using samples with a medium yeast lees addition of 20 g and 40 g. Statistically significant differences according to t-values were observed, especially for higher alcohol and acetate ester groups.
Figure 4. Antioxidant activity - FRAP method.
Figure 5. Antioxidant activity - DPPH method.
Table 2. T-test: higher alcohols.
| Average ± SD (basic) |
Average ± SD 20-g lees |
t | p | Average ± SD 40-g lees |
t | p | Average ± SD 80-g lees |
t | p | |
|---|---|---|---|---|---|---|---|---|---|---|
| Isoamyl alcohol | 103.42±1.35b | 88.10±3.94a | 6.38 | 0.00 | 128.66±4.19b | –9.20 | 0.00 | 139.34±4.51a | –12.39 | 0.00 |
| Isobutyl alcohol | 6.35±0.65a | 6.28±0.21a | 0.21 | 0.85 | 8.65±0.17b | –8.76 | 0.00 | 8.73±0.27a | –7.81 | 0.00 |
| 2-Phenylethanol | 5.49±0.15a | 5.35±0.26a | 0.85 | 0.45 | 9.12±0.42b | –12.59 | 0.00 | 8.22±0.53a | –7.81 | 0.00 |
| 1-Propanol | 3.36±0.19a | 3.06±0.07a | 2.57 | 0.06 | 4.39±0.05b | –8.27 | 0.00 | 4.65±0.16a | –8.50 | 0.00 |
| 1-Hexanol | 1.15±0.10b | 0.94±0.03a | 3.78 | 0.02 | 1.19±0.06a | –2.17 | 0.10 | 1.38±0.02a | –11.94 | 0.09 |
| 1-Butanol | 0.43±0.07a | 0.36±0.01a | 1.74 | 0.16 | 0.45±0.01a | –3.37 | 0.03 | 0.44±0.03a | –2.27 | 0.00 |
| Benzyl alcohol | 83.67±4.51a | 92.56±2.91b | –2.87 | 0.05 | 166.52±8.06b | –14.78 | 0.00 | 241.45±14.17a | –18.06 |
Note: Results are expressed as the mean value of three measurements ± standard deviation. The division into homogeneous groups (a,b) was based on Fisher’s test, the significance level is α = 0.005.
Table 3. T-test: acetate esters.
| Average ± SD (basic) |
Average± SD 20-g lees |
t | p | Average ± SD 40-g lees |
t | p | Average ± SD 80-g lees |
t | p | |
|---|---|---|---|---|---|---|---|---|---|---|
| Ethyl acetate | 28.28±1.14a | 28.12±1.18a | 0.17 | 0.88 | 25.22±1.31a | 2.52 | 0.07 | 28.22±0.78a | –0.07 | 0.95 |
| Isoamyl acetate | 241.66±6.03a | 322.52±12.85b | –9.86 | 0.00 | 193.52±11.57a | 4.89 | 0.01 | 156.21±13.84a | 8.17 | 0.00 |
| 1-Hexyl acetate | 10.00±1.00b | 3.52±0.11a | 11.17 | 0.00 | 0±0.00a | 226.13 | 0.00 | 0±0.00a | 226.13 | 0.00 |
| 2-Phenylethyl acetate | 24.33±0.58b | 14.28±0.39a | 25.03 | 0.00 | 19.84±0.34a | 21.11 | 0.00 | 13.76±0.39a | 41.93 | 0.00 |
| Isobutyl acetate | 6.67±0.58a | 10.97±0.55b | -9.38 | 0.00 | 6.46±0.28a | –1.36 | 0.25 | 6.97±0.38a | -2.80 | 0.05 |
Note: Results are expressed as the mean value of three measurements ± standard deviation.The division into homogeneous groups (a,b) was based on Fisher’s test, the significance level is α = 0.005.
Levels of acetate esters, such as isoamyl acetate, 1-hexyl acetate, 2-phenylethyl acetate, and isobutyl acetate, were determined on comparing base wine to variants with addition of yeast lees (Table 3). An increase in higher alcohols was observed in wines with higher doses of fermentation lees. According to t-values, the differences were statistically significant, especially for 2-phenylethyl acetate (responsible for honey notes in wine). A significant difference was observed for isoamyl acetate, responsible for aroma in bananas; its concentration increased to 80.85 mg L–1 during aging with yeast lees, as discovered by Bautista et al. (2007). A statistically significant difference was discovered for 1-hexyl acetate, responsible for fruity aroma, with 40-g yeast lees variant. This increase in 1-hexyl acetate could be due to the malolactic fermentation. The findings of Agouridis et al. (2008) and Tofalo et al. (2021) confirmed the ability of Oenococcus oeni bacteria, species of lactic acid bacteria, to produce higher alcohols.
The values for isoamyl acetate, 1-hexyl acetate, 2-phenylethyl acetate, and isobutyl acetate were determined on comparing base wine with 20-g yeast lees variant (Table 3). According to t-values, the differences were statistically significant, especially for 2-phenylethyl acetate (responsible for honey tones in wine).
Discussion
Based on the data obtained, it was found that fermentation yeast lees had a very positive effect on the amount of oxygen in wine because of their capacity to absorb dissolved oxygen and substantially increase antioxidant properties of wine. This fact was confirmed by multiple studies. For example, Comuzzo et al. (2015) compared the antioxidant properties of oenological products, including yeast lees, and determined that yeast lees had maximum ability to consume oxygen of the products tested. In addition, effectiveness of lees is enhanced by the amount of residual phenolic substances that significantly increase the rate of dissolved oxygen consumption. The present research also confirmed that sedimented lees contained a large number of adsorbed polyphenols; this increased the antioxidant properties of wine and the number of polyphenols with increase in the amount of added lees. The increase in total phenols observed after lees’ contact period could be due to a release of tyrosol and other amino acids with a phenolic -OH group. In addition to polyphenols, wines are enriched in several colloidal molecules, such as nitrogen compounds, polysaccharides, lipids, glucans, and mannoproteins (non-filamentous glycoproteins).
Bottled wines containing fermentation lees undergo significant changes in their aromatic profile during aging. We observed significant changes in higher alcohol and acetate ester groups. A similar conclusion was reached by Bautista et al. (2007). According to Perez-Serradilla and de Castro (2008), yeast lees in wines reduce the volatile content and thus the fruity aroma of wines.
The results explained the rate and degree of oxygen consumption by wines depending on whether they contained fermentation lees or were filtered before bottling. It is clear that fermentation lees have a major affect on the antioxidant properties of wine and can bind suspended oxygen more rapidly than the wine itself. Furthermore, lees protect the wine to a certain extent from possible chemical oxidation. However, this could not be applied to all wines. Light fruit wines of reductive style, and especially wines containing residual sugar, are not suitable for the use of this technology. Chemical composition, pH, technological operations, and storage temperature of a particular wine also play an important role.
Conclusion
The results of the study demonstrate that the addition of fermentation lees to bottled wine substantially accelerates the consumption of dissolved oxygen. This effect is due to the oxygen consumption of yeast lees. The rate of dissolved oxygen consumption depends on the dose of yeast lees. This addition also affects the chemical composition and certain parameters of wine, in particular the amount of reducing sugars, volatile acids and glycerol. We conclude that fermentation lees could bind dissolved oxygen at a higher rate than the wine itself. In addition, fermentation lees protect wines to a certain extent from possible chemical oxidation.
Author Contributions
Michaela Kulhankova and Bozena Prusova: conceptualization. Mojmir Baron: methodology and validation. Michaela Kulhankova: formal analysis, visualization and writing of original draft. Bozena Prusova: writing—review and editing. All authors read and agreed to the published version of the manuscript.
Funding
This paper was supported by the project CZ.02.1.01/0.0/0.0/16_017/0002334 Research Infrastructure for Young Scientists, and co-financed by the Operational Programme Research, Development and Education and the project Study of Oxygen in Various Technological Stages of Wine Production, IGA-ZF/2021-SI2008.
REFERENCES
Agouridis N., Kopsahelis N., Plessas S., Koutinas A.A. and Kanellaki M. 2008. Oenococcus oeni cells immobilized on delignified cellulosic material for malolactic fermentation of wine. Bioresource Technol. 99: 9017–9020. 10.1016/j.biortech.2008.04.026
Baron M. and Sochor J. 2013. Estimation of thiol compounds cysteine and homocysteine in sources of protein by means of electrochemical techniques. Int J Electrochem Sci. 8: 11072–11086. 10.1016/S1452-3981(23)13170-1
Bautista R., Fernández E. and Falqué E. 2007. Effect of the contact with fermentation-lees or commercial-lees on the volatile composition of white wines. Eur Food Res Technol. 224: 405–413. 10.1007/s00217-006-0336-7
Carmona-Jimenez Y., Garcia-Moreno M.V., Igartuburu J.M. and Garcia Barroso C. 2014. Simplification of the DPPH assay for estimating the antioxidant activity of wine and wine by-products. Food Chem. 165: 198–204. 10.1016/j.foodchem.2014.05.106
Comuzzo P., Battistutta F., Vendrame M., Páez M.S., Luisi G. and Zironi, R. 2015. Antioxidant properties of different products and additives in white wine. Food Chem. 168: 107–114. 10.1016/j.foodchem.2014.07.028
Day M., Schmidt S., Smith P. and Wilkes E. 2015. Use and impact of oxygen during winemaking. Aust J Grape Wine Res. 21: 693–704. 10.1111/ajgw.12199
Eisenman L. 1987. Oxygen uptake in wine. Enol Viticult. 5: 38.
Fornairon-Bonnefond C. and Salmon J.M. 2003. Impact of oxygen consumption by yeast lees on the autolysis phenomenon during simulation of wine aging on lees. J Agric Food Chem. 51: 2584–2590. 10.1021/jf0259819
Friedel M., Patz C.D. and Dietrich H. 2013. Comparison of different measurement techniques and variable selection methods for FT-MIR in wine analysis. Food Chem. 141: 4200–4207. 10.1016/j.foodchem.2013.06.120
Myers T.E. and Singleton V.L. 1979. The nonflavonoid phenolic fraction of wine and its analysis. Am J Enol Viticult. 30: 98–102. 10.5344/ajev.1979.30.2.98
Perez-Serradilla J.A. and de Castro M.D. 2008. Role of lees in wine production: a review. Food Chem. 111: 447–456. 10.1016/j.foodchem.2008.04.019
Prusova B. and Baron M. 2018. Effect of controlled micro-oxygenation on white wine. Ciência Técnica Vitivinícola. 33: 78–89. 10.1051/ctv/20183301078
Pulido R., Bravo L. and Saura-Calixto F. 2000. Antioxidant activity of dietary polyphenols as determined by a modified ferric reducing/antioxidant power assay. J Agric Food Chem. 48: 3396–3402. 10.1021/jf9913458
Salmon J.M., Fornairon-Bonnefond C. and Mazauric J.P. 2002. Interactions between wine lees and polyphenols: influence on oxygen consumption during simulation of wine aging. Food Sci. 67: 1604–1609. 10.1111/j.1365-2621.2002.tb08691.x
Schneider V., Muller J. and Schmidt D. 2016. Oxygen consumption by postfermentation wine yeast lees: factors affecting its rate and extent under oenological conditions. Food Technol Biotechnol. 54: 395–402. 10.17113/ftb.54.04.16.4651
Singleton V.L. and Rossi J.A. 1965. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am J Enol Viticult. 16: 144–158. 10.5344/ajev.1965.16.3.144
Sochor J., Jurikova T., Pohanka M., Skutkova H., Baron M., Tomaskova L., et al. 2014. Evaluation of antioxidant activity, polyphenolic compounds, amino acids and mineral elements of representative genotypes of Lonicera edulis. Molecules 19: 6504–6523. 10.3390/molecules19056504
Sochor J., Ryvolova M., Krystofova O., Salas P., Hubalek J., Adam V., et al. 2010. Fully automated spectrometric protocols for determination of antioxidant activity: advantages and disadvantages. Molecules 15: 8618–8640. 10.3390/molecules15128618
Sochorova L., Prusova B., Jurikova T., Mlcek J., Adamkova A., Baron M. et al. 2020. The study of antioxidant components in grape seeds. Molecules 25(16): 3736. 10.3390/molecules25163736
Tofalo R., Battistelli N., Perpetuini G., Valbonetti L., Rossetti A.P., Perla C., et al. 2021. Oenococcus oeni lifestyle modulates wine volatilome and malolactic fermentation outcome. Front Microbiol. 12: 736789. 10.3389/fmicb.2021.736789
Valade M., Tribaut-Sohier I., Bunner D., Pierlot C., Moncomble D. and Tusseau D. 2006. Les apports d’oxygene en vinification et leurs impacts sur les vins. Le Vigneron Champenois 8: 9.
Vidal J.-C. and Moutounet M. 2008. Control of the oxygen supply during the conditioning part 1: principles and wine preparation. Presented at the UFOE Congress Limoux, 30 May 2008. Revue Française d’Œnologie. N. 229 (Online). Available at: www.infowine.com
Vinventions. 2018. Wine Quality Solutions (WQS) NomaSense O2 P300 & P6000. Vinventions, Zebulon, NC. Available at: www.vinventions.com
Waterman P.G. and Mole S. 1994. Analysis of Phenolic Plant Metabolites. Blackwell Scientific, Oxford, UK.
WineBussines, 2019, Oct. Wine Quality Solutions Launches the NomaSense™ Oxymeter. WineBusiness, Sonoma, CA. Available at: www.winebusiness.com
Zhang P., Ma W., Meng Y., Zhang Y., Jin G. and Fang Z. 2021. Wine phenolic profile altered by yeast: mechanisms and influences. Compr Rev Food Sci Food Saf. 20: 3579–3619. 10.1111/1541-4337.12788
Zironi R., Comuzzo P., Tat L. and Scobiola S. 2010. Oxygen and wine. Internet J Enol Viticult. 3: 1–5.